
Retour au sommaire des cours |

Une version pdf est à votre disposition ici. |
v Introduction :
Les cellules neuronales :
Le neurone
possède 4 régions :
-
Le
corps cellulaire (ou soma) avec le noyau,
-
Les
dendrites,
-
Un
unique axone,
-
Les
terminaisons pré–synaptiques.
L’axone
est l’élément de transmission.
-
Sa
longueur est variable (peut atteindre le mètre), ainsi que son diamètre (0,2 à
20 µm par rapport au soma : 80 µm)
-
Il
y a la présence d’un cône d’implantation de l’axone (au niveau du soma), il
s’agit du segment initial (ou axon hillock) C’est lui qui produit les PA qui se
propagent vers l’axone.
-
Il
a la présence d’une gaine de myéline ou non, dont dépend la vitesse de
propagation du PA.
-
L’axone
se ramifie pour former de 10 à 1000 synapses.
Il existe 2
sortes de dendrites : les dendrites apicaux et les dendrites basaux. Il
s’agit de la surface majeure de réception.
Les cellules gliales :
Elles
sont 10 à 50 fois plus nombreuses que les neurones (chez les vertébrés)
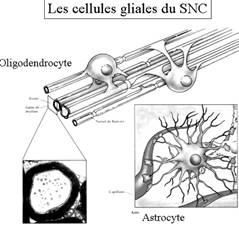

Leurs
rôles :
-
Elément
de support qui donne une certaine fermeté, stabilité, structure pour le
matériel cérébral.
-
Séparation
entre différents groupes neuronaux,
-
Production
de myéline (= manchon isolant) qui est assurée par 2 types de cellules
gliales :
-
Les
oligodendrocytes
(SNC) qui ont pour rôles la myélinisation de plusieurs neurones chacun
(dans la substance blanche) et l’enveloppement des corps cellulaires (dans la
substance grise)
-
Les cellules de
Schwann
(SNP) qui myélinisent une partie d’un axone.
Les astrocytes :
Ce sont les plus
nombreux dans la glie. Elles ont une forme étoilée et développent des pieds
astrocytaires sur les capillaires sanguins et les neurones.
Leurs
rôles :
-
Elimination
des débris après une lésion ou une destruction neuronale (« éboueur »),
-
Tamponnage
de la concentration extracellulaire en Ca2+,
-
Absorption
et traitement des transmetteurs chimiques libérés par les neurones lors de
transmission synaptique,
-
Pour certaines
classes d’astrocytes,
guide la croissance des neurones et dirige l’allongement des axones au cours du
développement,
-
Aide
à la formation d’un revêtement imperméable pour les capillaires : barrière
hémato–encéphalique (BHE) Comme toute barrière, il s’agit aussi d’une surface
d’échange,
-
Pour certaines, fonction
nutritive vers les neurones.

Le réflexe : du
latin refletere : réfléchir
C’est une
réaction motrice ou sécrétoire déclenchée par le système nerveux en dehors de
la volonté et en réponse à une stimulation des terminaisons des fibres
sensitives.
L’excitation
transmise par les fibres sensitives centripètes est réfléchie par une cellule
nerveuse centrale et renvoyée par une fibre centrifuge vers un organe plus ou
moins éloignée. La fibre centrifuge appartient soit au système nerveux
somatique, soit au système nerveux autonome.
Il
existe 2 types de réflexes :
-
Le réflexe
absolu (ou inconditionnel) :
Il est dit aussi
ordinaire ou congénital (= de l’espèce)
Il fait
intervenir un centre infra–cortical (comme le bulbe)
-
Le réflexe conditionné
(ou conditionnel) :
Il est dit aussi
acquis ou psychique.
Il
fait intervenir le cortex cérébral.
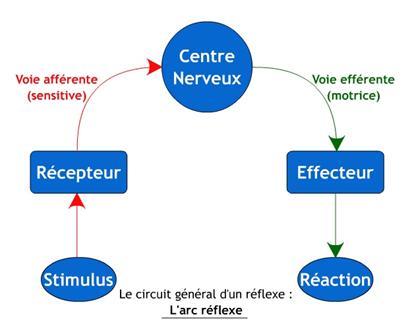
L’arc
réflexe :
C’est l’ensemble
neuronal mettant en jeu des récepteurs et des effecteurs liés entre eux par le
système nerveux central. Sa fonction est de contrôler en permanence les
réactions stéréotypées de l’organisme quand il est stimulé par l’environnement.
v La moelle épinière :
racine motrice et racine sensorielle :
Ø  Schéma neuronal de la moelle :
Schéma neuronal de la moelle :

Ø Expérience
de Magendie – expérience de dégénérescence :
§ Expérience de section :
La section des
racines dorsales lombo–sacrées (chez le chien) d’un coté de la moelle épinière.
" Il y a une anesthésie du membre postérieur de
ce coté, mais l’animal peut bouger sa patte.
La section des
racines ventrales de la même région.
" Toute stimulation de la patte est ressentie
par le chien, mais ce membre est complètement immobile.
§ Expérience de
stimulation :
Excitation du
bout périphérique dorsal :
" Aucune contraction musculaire n’est obtenue.
Excitation du
bout central dorsal :
" Mouvement réflexe et éventuellement une
réaction douloureuse.
Excitation du
bout central ventral :
" Aucune contraction musculaire n’est obtenue.
Excitation du
bout périphérique ventral :
" Contraction des muscles.
Þ
La voie dorsale comporte les informations sensitives venues des
récepteurs périphériques jusqu’au centre médullaires (SG) =
Influx sensitifs sensoriels
Þ
La racine ventrale transporte des information moteurs des centres
médullaires jusqu’aux effecteurs (comme les muscles)
Remarque :
« Sensitif »
pour les voies afférentes et « sensoriel » pour les organes
sensoriels.
§ Localisation des corps
cellulaires – expérience de dégénérescence :
La partie de fibre non reliée au soma
dégénère.
Section de la
racine dorsale entre le ganglion et la moelle :
" Dégénérescence entre le ganglion et la
moelle.
Section de la
racine dorsale entre le ganglion et le nerf périphérique :
" Dégénérescence du ganglion au nerf.
Section de la
racine ventrale au bon milieu :
" Dégénérescence du lieu de section au nerf.
Þ
Les somas des fibres efférentes se situent dans la substance grise = Cellule
motrice de la corne ventrale (ou motoneurones)
Þ
Les somas des fibres afférentes se situent dans les ganglions de la
racine dorsale (ou ganglion spinal)
Ø Niveau
supra–médullaire : aréflexie ou choc spinal :
Chez les
mammifères, et en particulier chez les primates, les structures nerveuses
situées au dessus de la moelle exerce un contrôle sur la moelle épinière et
donc inhibent le fonctionnement indépendant de la moelle.
Quand la moelle
est isolée du cerveau, elle est capable d’assurer certains réflexes et ce après
une phase d’aréflexie totale (= choc
spinal) qui est fonction du degré d’encéphalisation de l’animal : quelques
minutes chez un batracien, quelques mois chez l’homme.
L’aréflexie
est la conséquence de la suppression brutale des contrôles (facilitation et
inhibition) des diverses structures sus–jacentes (= supra–médullaires) sur la
moelle épinière.
De plus,
l’isolement par section de la moelle épinière des structures supra–médullaires
prive l’animal des fonctions élaborées dans ces régions, avec la disparition de
la motricité volontaire, la sensibilité et aussi les réflexes de complexité
plus grande que les réflexes médullaires.
La sensibilité citée ci–dessus est à
prendre au sens de sensibilité consciente :
-
La sensibilité
superficielle
(de la peau ou extéroceptive) correspond
aux stimulations tactiles classiques (température, douleur, pression, etc.)
Elle
utilise la voie spino–thalamique (ou voie extra–lemniscale)
-
La sensibilité profonde (ou
proprioceptive) correspond aux
stimulations musculaires, tendineuses, osseuses ou articulaires.
v Réflexe monosynaptique
(exemple du réflexe myotatique ou réflexe d’étirement) :
Ø Définition
du réflexe :
Exemples : Lors d’une
brûlure, le réflexe cornéen (clignement des yeux) ou le réflexe d’atténuation
au niveau de l’audition.
Ce
dernier a un rôle dans la transmission du son de l’oreille interne. Il met en
jeu le muscle tenseur du tympan et le muscle stapedius (entre la paroi osseuse
et l’étrille = 3ème osselet)
Il consiste en la contraction de ces 2 muscles. La chaîne des osselets devient
plus rigide et les sons se transmettent moins bien vers le volet ovale.
L’atténuation
est plus marquée pour les basses fréquences que les hautes fréquences. Ce
réflexe intervient que 50 à 100 ms après l’arrivée du son. Il ne protège donc
pas des sons violents.
Ø Fuseau
neuromusculaire = récepteur de l’arc monosynaptique du réflexe
d’extension :
Le réflexe
myotatique a été découvert par Lidder et Sherrington.
Il consiste en
la contraction d’un muscle quand il est étiré : l’étirement du muscle
stimule les récepteurs de ce muscle.
Exemple : Un coup de
marteau réflexe sur le tendon du genou entraîne l’extension du muscle
quadriceps.
C’est
un réflexe propriocepteur qui permet le maintien du tonus musculaire (posture,
exercices) et protège d’un étirement excessif du muscle.
L’étirement
est constitutif à un mouvement articulaire, au poids du corps ou à la
contraction des muscles antagonistes.
Un
propriocepteur est un récepteur
situé dans les muscles, les tendons et les articulations, et qui fournit des
informations au SNC sur la position et le mouvement du corps.
La proprioception
est la réception des informations venant des muscles, tendons, articulations et
également du labyrinthe (partie de l’oreille interne pour l’équilibration) On
parle aussi de kinesthésie (kinesis =
mouvement et aïsthesis = sensibilité)
ou encore de « sens musculaire ».
§ Fuseau neuromusculaire :
Il
s’agit de fibres musculaires de diamètre plus faible (15 – 30 µm) et plus
courtes (4 –
Les fibres
extrafusales (très longues) vont d’un tendon à l’autre.
Terminaisons motrices g Axones efférents Axones afférents Terminaisons sensorielles Capsule Fibres musculaires
intrafusales

Innervation sensorielle du fuseau
neuromusculaire :
Il
s’agit des fibres afférentes au SNC qui s’enroulent autour des fibres centrales
du fuseau pour former une terminaison annulo–spirale. Ce sont des fibres
grosses et myélinisées (= fibres Ia)
La
région musculaire au centre est un peu bombée et ne se contracte pas. Si les
fibres des régions distales se contractent, cela entraîne un étirement de la
région médiane.
" Plus cette
région est étirée, plus la fréquence des PA des fibres Ia est augmentée.
Innervation motrice :
-
Pour
les fibres extrafusales, il s’agit des fibres a ou Aa issues des
motoneurones a (de la corne ventrale)
-
Pour
les fibres intrafusales, il s’agit des fibres g ou Ag dont les corps cellulaires sont
présents dans la moelle épinière.
Les fibres a ont un plus
grand diamètre que les fibres g.
Fonctionnement :
La membrane des
terminaisons sensorielles présente des canaux ioniques sensibles à l’étirement (stretch
channels), au niveau de la membrane nerveuse, et reliés au muscle par des spectrines.
Il s’agit de
canaux sélectifs aux cations Na+ et très
probablement Ca2+. La
driving–force (Vm–Eion) pousse vers une entrée de cation, créant une
dépolarisation. Il s’agit du potentiel de récepteur. Il peut y avoir la
somation des potentiels de récepteur.
Pour les
expériences, on utilise du TTX pour inhiber les PA qui se mélangeraient aux
potentiels de récepteurs.
-
La
dépolarisation du fuseau neuromusculaire, en réponse au changement de longueur
du muscle (réponse dynamique), est proportionnelle à la fois à la vitesse et à
l’amplitude de l’étirement.
-
Lorsque
l’étirement est maintenu à une longueur fixe, le potentiel de récepteur diminue
vers une valeur plus faible. Il n’est plus proportionnel qu’à l’amplitude de
l’étirement.
-
Au
repos, les canaux sensibles à l’étirement s’ouvrent sporadiquement pendant des
intervalles de temps courts, ce qui produit un courant dépolarisant
transitoire.
-
Lorsque
la pression négative sur la membrane augmente (succion), le canal s’ouvre plus
souvent et reste à l’état ouvert pendant des intervalles de temps plus longs.
Notion de terminaison primaire et
terminaison secondaire :
Fibres à chaîne Fibre à sac statique Fibre à sac dynamique Fibre motrice g dynamique II Ia Fibre motrice g statique

Les fibres à sac
présentent un renflement ; les fibres à chaîne non.
Quand
un muscle est étiré ou relâché, à partir de l’étirement initial, il y a 2
phases de variation de longueur :
-
Une
phase dynamique qui correspond au changement de longueur du muscle,
-
Un
phase statique (ou état stable) où le muscle est stabilisé dans sa nouvelle
longueur.
Travaux de
Cooper :
Pendant
la phase dynamique d’étirement, les terminaisons IAIRES ont une
activité de fréquence plus élevée que pendant la phase d’état stable
ultérieure.
Les terminaisons IIAIRES ont une
augmentation de leur activité de fréquence de manière graduelle et n’est pas
beaucoup plus élevée pendant la phase dynamique que pendant la phase d’état
stable ultérieure atteinte.
Les
terminaisons IAIRES du fuseau
neuromusculaire sont hautement sensibles à la vitesse de changement (ou
variation) de longueur du muscle.
" L’augmentation
de la fréquence d’activité de ces terminaisons reflète la vitesse de variation
de longueur du muscle.
Pendant un
raccourcissement rapide, les terminaisons IIAIRES font une pause
puis reprennent leur activité à une fréquence plus basse lorsque le
raccourcissement s’arrête.
Les 2 types de
terminaisons :
Etirement linéaire
Coup léger
Vibration
Relâchement
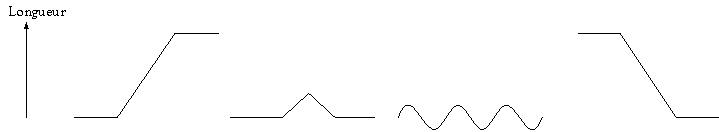


Pour les stimuli
transitoires (coup léger ou étirement bref) ou lorsque le muscle est soumis à
des vibrations, cela produit des salves (ou bursts) de PA donc des activités
(ou firings) des terminaisons IAIRES. Elles ont une
sensibilité qui augmente avec le changement de longueur, surtout lors de la
phase dynamique.
Au contraire,
les stimuli brefs n’ont pratiquement pas d’effet sur les terminaisons IIAIRES. Ces variations
de longueur surviennent trop rapidement pour modifier la décharge de l’état
stable d’une terminaison IIAIRE.
Les
terminaisons IAIRES codent non
seulement la longueur du muscle et ses variations, mais aussi la vitesse de
variation de longueur.
" Information sur la vitesse de mouvement et de
la position statique.
Lors d’un
relâchement, il y a une absence de PA : il est donc aussi signalé.
Les petites variations du muscle (inf. à
Remarque :
La
sensibilité dynamique des terminaisons IAIRES diminue quand
il y a de grandes variations de longueur. Ces terminaisons sont capables de
retrouver leur aptitude à répondre à de petites variations de longueur, même
quand elles ont atteint une nouvelle longueur elles–même.
" Elles sont capables de détecter (to sense) de petites variations de
longueur quelque soit la longueur d’état stable du muscle.

6
mV
0
Réponse dynamique
Réponse d’état stable
§ Expérience d’étirement du
muscle = réflexe monosynaptique d’étirement :
Inhibition antagoniste :
+
Elle
ne correspond pas à un réflexe myotatique. Les fibres IA constituent des
synapses excitatrices sur les motoneurones homonymes (du même muscle) et des
synapses inhibitrices sur les motoneurones antagonistes.
– – + + +![]()
![]()
![]()
Fibres
Ia Moelle
épinière Motoneurone
Réflexe myotatique sans inhibition
antagoniste :


Contraction (= réponse réflexe)
Etirement
Lors d’un bref
étirement, il y a génération de PA qui parcourt les fibres Ia des terminaisons IAIRES vers la moelle
épinière et qui engendre un PPSE.
PPSE = Potentiel Post–Synaptique Excitateur
= Mauvaise traduction de l’anglais de
EPSI = Excitatory Post–Synaptic Potential
Il faut préférer cette version pour
mieux concevoir un PPSE (signification réelle)
=
End–Plate Synaptic Potential (end–plate
= plaque motrice)
Le potentiel
passe de –70 mV (potentiel de repos) à –60, –55 mV.
Quelques uns de
ces PPSE sont supra–linéaires et permettent la génération d’un PA.
Réflexe myotatique généralisé :

Il y a
l’intervention d’un interneurone inhibiteur sur la voie antagoniste.
Ø Les
diverses parties d’un arc réflexe :
§ Nomenclature générale :

![]()

![]()

§ Arc du réflexe monosynaptique
d’étirement – temps réflexe :
Les effecteurs
et les récepteurs sont inclus dans un organe unique : le muscle
squelettique. On parle aussi de réflexe intrasèque.
Le
délai synaptique est de 0,2 à 0,5 ms mais il peut être beaucoup moins long.
Le temps réflexe
est le délai entre l’instant du stimulus et la réponse de l’effecteur.
Exemple : le réflexe rotulien.
Ce délai se
compose :
-
Du
temps de propagation des PA afférents (fibres Ia),
-
Et
du temps de propagation des PA efférents (axone moteur a)
La
vitesse de propagation des PA dans ces fibres est de
t =
d/v = 1,6/100
= 1,6 . 10–2 = 16
ms
A
ce temps, il faut ajouter 9 à 14 ms pour les évènements suivants :
-
Délai
entre le début de l’étirement et le 1er PA au niveau du fuseau
neuromusculaire,
-
Transmission
au niveau des synapses des motoneurones : délai synaptique (soit 0,2 à 0,5
ms) pour la plupart des synapses périphériques ou centrales,
-
Transmission
des informations des plaques motrices aux fibres musculaires (dépolarisation
électrotonique locale déclenchant un PA = réponse propagée et régénérative),
-
Propagation
du PA le long de la fibre musculaire,
-
Induction
de la contraction par le PA : couplage excitation – contraction (ou
couplage électromécanique)
Þ D’où un temps de réflexe monosynaptique
d’étirement de 25 à 30 ms.
Le temps réflexe
est différent du délai synaptique.
§ Fibres intrafusales et fibres
extrafusales : Fonction :
Activation d’une terminaison sensorielle
IAIRE :
Elle
est due à l’étirement passif d’un muscle allongé (avec les fibres intra– et
extrafusales)
Il est possible
d’exciter les terminaisons IAIRE des fuseaux
neuromusculaires en provoquant la contraction des fibres neuromusculaires
intrafusales par une stimulation arrivant par les fibres g (motrices)
La contraction
des fibres intrafusales ne changent ni la longueur ni la tension (force)
exercée sur l’ensemble des fibres musculaires, car cette contraction exerce une
force trop faible et ce même si toutes les fibres intrafusales se contractent.
La
stimulation g entraîne la
contraction des régions polaires des fibres intrafusales et le raccourcissement
de ces régions. La région centrale reste allongée.
Cela entraîne la production de PA dans
les terminaisons IAIRES (dans les
fibres Ia) comme dans le cas d’étirement du muscle en entier. On observe
ensuite la formation de PPSE sur le motoneurone a (fibre
extrafusale)
 Etirement du muscle et contraction des
fibres intrafusales : effet complémentaire ou antagoniste :
Etirement du muscle et contraction des
fibres intrafusales : effet complémentaire ou antagoniste :

j Contraction des fibres intrafusales (g)
+ Etirement des muscles

k Contraction des fibres extrafusales (a)
+ Relâchement fibres intrafusales (g)

l
Modulation
de la contraction des fibres intrafusales (a) entre contraction
maximum et relaxation
Le seuil
d’excitation des récepteurs peut être modifié par une mise sous tension
préalable des fibres intrafusales.
Voie motrice :
-
Les
motoneurones a sont sous contrôle
cervical pour la voie du faisceau pyramidal.
-
Les
motoneurones g sont sous
contrôle du tronc cérébral avec en particulier la substance réticulée (ou
formation réticulée) Ces régions sont en relation avec le cervelet, les noyaux
gris centraux et avec le cortex moteur.
Les
noyaux gris centraux, appelés aussi ganglions de la base, sont
constitués :
-
Du
putamen,
-
Du
pallidum (ou globus pallidus),
-
Du
noyau caudé,
-
De
la substance noire (ou substantia nigra
ou locus niger),
-
Du
noyau sub–thalamique (ou noyau sous–thalamique
ou corps de Luys)
Adaptation des récepteurs :
L’adaptation
est soit rapide, soit lente.
Les
mécanorécepteurs à adaptation rapide (dans les articulations) fournissent une
information dynamique concernant la position du membre et le niveau d’articulation.

v Notion de réflexe moteur
polysynaptique :
Ø Définition :
Dans
les arcs réflexes moteurs polysynaptiques, plusieurs neurones sont reliés en
série (interneurone) et le motoneurone n’est que le dernier de la chaîne.
En général, le récepteur et l’effecteur
sont éloignés : Réflexe extrasèques.
Cela permet un ajustement plus facile de la réponse réflexe aux nécessités de
l’organisme (locomotion, alimentation, protection du corps dans l’environnement
= réflexe de défense)
Ø Exemples
de réflexes polysynaptiques :
§ Quelques réflexes :
Réflexe de succion :
Lorsque
que le sein est présenté à la bouche, cela entraîne un réflexe de succion de la
part du bébé. Ce réflexe peut être obtenu en touchant les lèvres avec le doigt.
-
Les
récepteurs sont les mécanorécepteurs de la peau des lèvres (mais aussi les
thermorécepteurs),
-
Les
effecteurs sont les muscles des lèvres, des joues, de la langue, du thorax et
du diaphragme.
Ce réflexe est
très complexe : il doit coordonner la respiration avec le passage du lait
dans l’œsophage (et non dans la trachée)
Réflexe de défense :
Une
grenouille décérébrée peut rester en vie plusieurs heures.
On met de
l’acide (juste assez concentré pour causer une démangeaison) sur du papier
filtre que l’on pose dur le dos de la grenouille. On observe un mouvement
typique avec la patte postérieure la plus proche du papier pour enlever le
papier.
-
Les
récepteurs sont les nocicepteurs de la peau de dos.
-
Les
effecteurs sont les muscles de la patte postérieure.
La contraction
musculaire a une apparence intentionnelle, volontaire et bien coordonnée malgré
l’absence du cerveau.
La coordination
est telle que la réponse est différente selon la posture relative des membres.
Cela démontre une adaptation très fine du mouvement. On parle de pattern,
motif, séquence (voire de patron)
§ Caractéristiques des réflexes
polysynaptiques :
Cela concerne le
temps réflexe, l’intensité et l’extension du réflexe.
Réflexe de la toux :
Il
permet la libération de passage dans la trachée (= réflexe de défense) Les
effecteurs sont les muqueuses de la trachée et des bronches.
Un chatouillement ou une irritation de
la gorge n’entraînent pas une toux immédiate, mais après un petit délai.
Les stimuli
infraliminaires se somment et forme un stimulus supraliminaire à condition
qu’ils durent assez longtemps. Cette sommation est un phénomène central, dans
les interneurones des arcs réflexe (et non dans les récepteurs périphériques)
Si l’intensité
augmente, le temps réflexe diminue. Cependant une excitation supraliminaire
plus vite atteinte dans les neurones centraux de l’arc réflexe tient aussi un
grand nombre de récepteurs plus intensément excités.
La baisse du
temps réflexe est surtout dû à des facilitations temporelles et spatiales.
 Facilitation temporelle et spatiale :
Facilitation temporelle et spatiale :
La
production répétitive des PPSE a un effet excitateur additif sur le neurone.
La
facilitation temporelle consiste en l’augmentation de l’excitabilité sur le
PPSE.
Les
décharges des récepteurs périphériques se produisent répétitivement, s’ajoutent
et produisent au niveau des synapses des neurones centraux des PPSE
supraliminaires

C’est une
différence spatiale qui entraîne une différence temporelle (temps d’arrivée du
signal au soma)
Facilitation temporelle et spatiale :
Légende : Dépolarisation infraliminaire Dépolarisation supraliminaire

Þ 3 Þ 2 Þ 8 > 3+2
=
Facilitation spatiale
Phénomène d’occlusion :

Þ 6 Þ 6 Þ 8 < 6+6
=
Phénomène d’occlusion
Ø Comparaison
réflexe moteur – réflexe viscéraux :
§ Réflexe autonome :


§ Réflexe polysynaptique
mixte :
Il fait partie à
la fois au SNS et au SNA.
Le réflexe de la toux :
C’est un réflexe
de défense présentant des récepteurs dans les muqueuses trachéennes et
bronchique (= viscérorécepteurs)
Des réflexes
complexes possèdent des voies afférentes à la fois des fibres du SNS et aussi
des fibres du SNA (autre exemple : les réflexes sexuels)
La
plupart des motoneurones et interneurones participent au fonctionnement de
plusieurs arcs réflexe.
Les axones moteurs de la gorge participent
à la déglutition, le réflexe de succion, le réflexe de toux, le réflexe
d’éternuement, et le réflexe respiratoire.
Ø Réflexe
de miction chez l’homme adulte :
La miction
permet la vidange du réservoir urinaire. Elle dépend de nombreux arcs réflexes.
La vessie est
une poche contractile, sa paroi est constituée de muscles lisses. Il y a 2
sphincters :
-
Le sphincter
externe strié
(sous le contrôle du système nerveux cérébro–spinal) qui maîtrise volontairement
le début et la fin de la miction.
-
Le sphincter
lisse
(interne) qui effectue la commande
automatique de la miction. C’est le seul en jeu chez l’enfant ou chez un malade
porteur d’une section spinal (de la moelle)
Vidange volontaire :
Elle est sous la
dépendance du centre supérieure cérébro–spinal. Elle est accompagnée de la
contraction du diaphragme et des muscles abdominaux, ainsi que la fermeture des
cordes vocales.
Mécanisme automatique :
Il
est sous la dépendance des centres lombo–sacrés (le centre sympathique lombaire
et le centre parasympathique sacré)
-
Le
système parasympathique assure la contraction du détrusor (= muscle vésical) et l’ouverture du sphincter lisse.
-
Le
système sympathique assure le relâchement du détrusor et la fermerture du
sphincter lisse.
Les récepteurs
principaux de la paroi, présents dans la vésicule et le sphincter lisse, sont
des terminaisons encapsulées (sensibles à la pression, au froid, à la traction)
Il existe aussi des terminaisons libres
nociceptives (= sensibles à la douleur)
Les voies afférentes
passent par le plexus hypogastrique
inférieur.
Les récepteurs
du sphincter strié sont des fuseaux neuromusculaires dont l’innervation
sensitive empreinte le nerf honteux
vers la corne dorsale de la moelle épinière en S2.
-
Les
récepteurs viscéraux sont responsables de la sensation de réplétion.
-
Les
récepteurs sphinctériens sont responsables de la sensation de besoin de
miction.
Les
centres segmentaires (= de la moelle épinière) de la miction regroupent :
-
Le centre
parasympathique sacré (S2, S3 et S4)
Sa destruction
entraîne une vessie automatique, dont la contraction est initiée par les plexus
intramuraux.
Plexus : ensemble de fibres nerveuses
entre–croisées (reliées entre–elles)
-
Le centre
sympathique thoraco–lombaire (de T12 à L2)
Il
inhibe le détrusor (= relâchement) et stimule le sphincter interne
(contraction)
Sa destruction
peut entraîner des rétentions.
-
Le centre cérébro–spinal
(C4)
Il commande le sphincter strié.
-
Les centres sus–jacents
Ils sont informés
de l’état de la vessie par les voies spino–thalamiques. Dans la formation
réticulée, il existe des couples de centres :
-
Aire
vésico–constrictive
+ Aire vésico–relaxante dans
l’encéphale,
-
Aire
vésico–constrictive
+ Aire sphinctéro–relaxante
(sphincter externe) dans le pont,
-
Aire
vésico–constrictive
+ Aire vésico–relaxante dans le
bulbe.
-
Il
existe aussi une interconnexion avec les centres de la respiration avec la
présence d’un phénomène d’apnée au début de la miction.
-
Les
fibres descendantes ont une action sur :
-
Les centres spinaux sous–jacents (C3 à T12)
responsables la pression intra–abdominale.
-
Les centres autonomes thoraco–lombaires de
la miction.
Le cortex
cérébral est impliqué par le bulbe avec :
-
La partie
pariétale (aires 1, 2 et 3) " Sensation vésico–urétrale.
-
La partie
frontale (aires 4 et 6) " Commande volontaire du sphincter strié.
-
Le gyrus (ou
circonvolution) cingulaire du cortex
cérébral " Partie antérieure.
-
Les fibres
cortico–spinales " Action sur le cortex (commande pour arrêter ou
initier la miction)
Conclusion :
La
miction est dépendante de 3 niveaux :
-
Le
niveau spinal autonome (siège des réflexes)
-
Le
niveau supra–spinal réticulaire (siège d’automatisme) responsable de la
coordination des autres actions concomitantes.
-
Le
niveau conscient (cortex)
Cette
disposition et les mécanismes expliquent que la lésion spinale au dessus du
renflement lombaire permet la récupération de la miction réflexe
quasi–automatique par les centres sympathique et parasympathique.
Si un problème
survient au niveau du renflement lombo–sacré, cela entraîne des désordres
majeures au niveau de la miction (rétention ou incontinence totale)
Ø Réflexe
inné et réflexe acquis :
§ Réflexe inné :
Il
s’agit de réactions stéréotypées de l’organisme et prédéterminées dans
l’organisation structurale du SNC.
Le réflexe inné
présente une forme quasi–identique chez tous les individus d’une même espèce.
Les neurones assurant le fonctionnement des arcs réflexe innés appartiennent
pour la plupart aux régions phylogénétiques les plus anciennes du SNC (moelle
épinière et tronc cérébral pour ce que
l’on vient de voir)
§ Réflexe acquis :
Tout individu
possède, en plus, la possibilité d’acquérir des réactions réflexe qui
permettent à son organisme de mieux répondre aux situations continuellement
changeantes de son environnement.
Les
arcs réflexes des réactions acquis mettent en jeu les régions supérieures du
SNC.
Les réflexes
peuvent aussi être oubliés et les possibilités d’acquisition sont contraintes
(dépendantes) de l’appartenance de l’individu donné à une espèce particulière.
= Réflexe
conditionnel = Conditionnement opérant
Hyporéflexie des réflexes
superficiels :
Elle consiste en
la baisse de la rigueur des réflexes superficiels (= de la peau)
Exemples
de réflexes superficiels :
-
Le
réflexe cornéen,
-
Le
réflexe cutané abdominal (= Contraction des muscles superficiels de l’abdomen),
-
Le
réflexe crémastérien (chez l’homme)
Il
consiste en l’élévation du scrotum après excitation de l’intérieur de la
cuisse.
v Notion générale sur la
motricité :
Ø Définition
hiérarchique des centres moteurs :
La
motricité est l’ensemble des mécanismes permettant à l’organisme :
-
De
mouvoir le corps et les membres par rapport aux objets environnants
" Contrôle des mouvements
-
De
maintenir une posture (attitude du corps dans l’espace)
" Contrôle postural
Les interactions
motrices avec l’environnement se font grâce aux muscles squelettiques
(exemples : communication orale, écriture, langage gestuel, chant)
Les
centres moteurs sont localisés à différents niveaux du système nerveux. Ces
structures sont organisées selon un plan hiérarchique : depuis les
structures les plus anciennes phylogénétiquement (la moelle épinière) aux plus
récentes (le cortex)
L’évolution n’a presque pas réalisé de remodelage
des structures existantes mais plutôt des superpositions à ce qui existait
déjà.
Ø Les
éléments constitutifs de cette hiérarchie :
§ La moelle épinière :
On y retrouve
les motoneurones a à l’origine des nerfs moteurs.
Ils sont appelés voie finale commune
car c’est là que converge toutes les informations du SNC.
§ Le tronc cérébral :
Il est chargé de
l’intégration des commandes motrices descendantes et des informations
sensitives ascendantes. Il est aussi le sélecteur des différentes catégories
d’informations.
Intégration synaptique :
La plupart des
neurones reçoivent plus ou moins simultanément des milliers d’informations
synaptiques qui activent différentes combinaisons de récepteurs canaux et de
récepteurs associés à des protéines G.
Le neurone post–synaptique intègre tous
ces signaux ioniques et biochimiques et génère un signal simple (PA) ou un
train de PA. La transformation de nombreux influx synaptiques en un seul type
de signal est à la base de l’intégration de l’information neuronale.
Le cerveau peut
aller jusqu’à des milliards d’opération par seconde.
§ Le cervelet :
C’est un comparateur. Il est capable de comparer
le programme moteur avec le mouvement réellement réalisé. Il est aussi capable
d’ajustements correctifs pendant le mouvement lui–même.
§ Le cortex moteur et
associatif :
Le cortex moteur
IAIRE se situe surtout dans l’aire 4.
On y trouve l’origine principale du faisceau pyramidale (descendant) qui forme
la voie pyramidale (ou voie motrice volontaire)
Le cortex moteur
associatif se situe dans le cortex frontal prémoteur puis dans le cortex
pariétal. Il est responsable de l’identification de la cible, du choix du
trajet, de la coordination sensorimotrice et la programmation du mouvement.
Le cortex agit
principalement par l’intermédiaire de l’aire 4 et présente des connexions
directes avec les structures sous–jacentes.
Ø 4
caractéristiques de cette organisation hiérarchique :
j Chaque composant du SNC comporte des cartes
somatotopiques. Le cortex moteur IAIRE possède une
représentation topographique avec les différents muscles du cortex.
Par exemple : l’aire motrice IAIRE est une zone
corticale dont les stimulations électriques nécessitent le moins d’énergie pour
générer un mouvement. Cette zone contient des cellules de Betz (= cellules
pyramidales géantes) On appelle aussi ce cortex : le cortex pyramidal.
La somatotopie
se voit sur l’homoncule de Penfield. On y observe 2 grandes zones au niveau du
cortex moteur : le visage et la main (meilleure représentation)
k Chaque niveau cortical reçoit une information
du cortex moteur provenant d’un périphérique (rétroaction) et est donc informer
de ses propres actions.
Cela permet une
adaptation et correction de son fonctionnement (avec l’aide du cervelet)
l Les différents niveaux du contrôle moteur
sont également organisés en parallèle de telle sorte que chacun d’entre–eux
(donc tous simultanément) soit capable d’agir directement sur le système
médullaire (c’est–à–dire sur les motoneurones a)
Hiérarchie du
contrôle nerveux de la motricité :
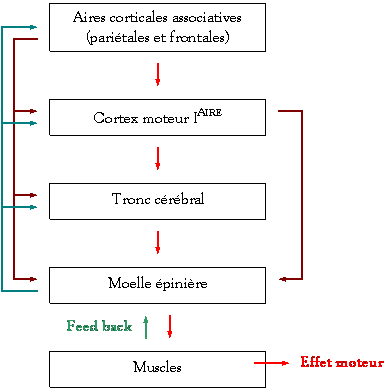
Þ
m Les autres structures qui interviennent dans
la motricité sont :
-
les
noyaux gris centraux (ou ganglions de la base)
-
Le
cervelet.
Elles modulent
l’activité de la hiérarchie des centres moteurs.
-
Les
aires associatives réalisent les fonctions les plus complexes :
apprentissage (cortex préfrontal), interprétation sensitive (aire gnostique),
le langage (aires de Broca = 44 et 45 + aire de Wernicke = aire 22)
Gnosie (gnosis = connaissance) = faculté
permettant de reconnaître par l’un des sens
la forme d’un objet, de se le
représenter et d’en saisir la signification
-
Le
striatum (ou corps striés ou encore corpus striatum) est une partie du
télencéphale qui constitue les noyaux profonds télencéphaliques. On y
trouve :
-
L’archistriatum
qui forme le noyau amygdalien,
-
Le
paléostriatum qui forme le globus pallidus et
-
Le
néostriatum qui forme le noyau caudé et le putamen (qui font partie des noyaux
gris centraux)
Ø Aires
corticales motrices : fonctions motrices du cortex (ainsi que du cervelet
et des ganglions de la base :
En ce qui
concerne les mouvements volontaires
§ Le cortex moteur IAIRE :
Les neurones de
cortex moteur IAIRE codent la
force, la direction et le sens des mouvements volontaires. Ils sont informés
des conséquences du mouvement et modifient leur activité pour anticiper les
mouvements.
§ Les aires corticales
prémotrices :
Elles préparent
le système moteur pour le mouvement.
-
Le
cortex prémoteur contrôle les
mouvements proximaux qui se projettent les bras vers une cible (par exemple)
-
Les aires motrices supplémentaires
programment (élaborent) les séquences motrices et coordonne les mouvements bilatéraux.
Elles permettent de réaliser mentalement une tâche sans l’exécuter de façon
motrice (abstraction)
§ Le cervelet :
Il
possède le cortex cérébelleux qui est divisé en 3 parties distincts. Il
participe à l’apprentissage moteur.
Les pathologies
du cervelet consistent
en des désordres
de la coordination.
§ Les ganglions de la base =
les noyaux gris centraux :
Ils
intègrent les informations en provenance des différentes aires du cortex. Ils
sont composés des 5 noyaux sous–corticaux :
-
Le
noyau caudé,
-
Le
putamen,
-
Le
globus pallidus (composé d’un segment interne et d’un segment externe),
-
Le
noyau sub–thalamique,
-
La
substancia nigra.
Ils
reçoivent des entrées en provenance du cortex, les intègrent et projettent en
retour vers le cortex et le thalamus.
La perte de
cellules dopaminergiques dans la
substancia nigra
provoque la maladie de Parkingson.
§ Le cortex prémoteur :
Il
est constitué de l’aire motrice supplémentaire et de l’aire prémotrice
(localisation différente)
La
stimulation des aires prémotrices entraîne une contraction coordonnée des
muscles au niveau de plus d’une articulation et, dans le cas d’une aire motrice
supérieure, des 2 cotés du corps.
Ces mouvements sont complexes et
nécessitent des courants stimulants de grande amplitude que les mouvements
produits par une stimulation du cortex moteur IAIRE.
-
Une
lésion du cortex moteur IAIRE cause un
affaiblissement des réponses motrices.
-
Une
lésion des aires prémotrices détériore l’aptitude à développer une stratégie de
mouvements appropriés.
Expérience :
Des singes sont
porteurs de lésions des aires prémotrices. On leur présente de la nourriture
derrière une vitre présentant une ouverture à coté.
" Les singes essaient de saisir directement la
nourriture, en se cognant contre la vitre, sans chercher à la contourner.
·
L’aire
motrice supplémentaire :
Les symptômes
chez les singes sont similaires aux cas d’apraxie qui surviennent chez l’homme
porteur de lésions des régions cervicales motrices supplémentaires (ou
d’associations pariétales supérieures)
Apraxie (praxis = action) = impossibilité de
conformer le mouvement au but proposé.
Ces patients ne
présentent ni affaiblissement, ni perte de sensitivité, et sont capables de
réaliser des mouvements simples avec précision. Mais ils ne sont pas capables
de réaliser des mouvements complexes nécessitant des séquences de contractions
de muscles, de réaliser une stratégie et de planifier des mouvements (comme se
brosser les dents)
§ Les aires associatives :
Le cortex peut
être divisé en aires sensitives (ou sensorielles) IIAIRES et en aires
motrices IAIRES. On parle alors
de cortex sensori–moteur. Le reste correspond au cortex associatif qui s’est
développé pendant l’évolution (dont l’aire associative frontale qui est
importante)
De
plus, les aires associatives semblent former des ponts (liaisons) entre les
cortex dévolus aux différentes modalités sensorielles, ainsi qu’entre le cortex
sensitif et le cortex moteur IAIRE.
Au
centre, se trouve le cortex pariétal postérieur (= aires 5, 7, 39 et 40 de
Brodmann) on peut s’attendre à une implication dans la coordination des
informations visuelles, auditives, somato–sensorielles, et les aires motrices
qui les entourent.
|
Fonction : |
Localisation : |
Site cortical : |
|
Aires
sensorielles IAIRES |
||
|
Visuelle |
Occipitale |
|
|
Auditive |
Temporale |
|
|
Somesthésique |
Pariétale |
Circonvolution
postérieure |
|
Aires
sensorielles associatives (IIAIRES) |
||
|
Somesthésique |
Pariétale |
Pariétal
postérieur (aires 5 et 7) |
|
Aires
associatives multimodales |
||
|
Intégration
sensorielle postérieure (localisation
visio–spatiale, attention, langage) |
Pariéto–temporale |
Jonction entre
les lobes (aires 7, 39
et 57) |
|
Intégration
motrice antérieure (planification
motrice, production du langage, jugement) |
Frontale |
Zone
préfrontale en avant des aires prémotrices (aires 45 et
46) |
|
Limbique (émotionnel,
mémoire) |
Tempo–pariétal |
« système
limbique » =
circonvolution cingulaire (aires 23 et
24) |
|
Aires
motrices associatives |
||
|
Région
prémotrice (programmation
et préparation motrice) |
frontale |
En avant de
l’aire motrice IAIRE |
|
Aires
motrices IAIRES
|
||
|
Cortex moteur (commande
d’excécution motrice) |
Frontale |
Circonvolution
pré–rolandique |
v Les systèmes moteurs
médullaires :
Les centres du SNC
impliqués dans la posture et les mouvements ne peuvent atteindre leur
planification que s’ils reçoivent un flux continuel sensitif. On parle de
système sensori–moteur. Les 2 organes associés au muscle squelettique sont le
fuseau neuromusculaire et l’organe tendineux de Golgi.
Ø L’organe
tendineux de Golgi :
§ Relation des composants de
base dans le muscle squelettique :

Les
organes tendineux de Golgi sont en série avec les fibres extrafusales.
Diamètre des
fibres : Ia ~ a >
Ib > II
> b

§ L’organe tendineux de
Golgi :

Il y a un
organe tendineux de Golgi pour 10 à 15 fibres intrafusales. Les organes
tendineux de Golgi est sensible à la force de traction du muscle, qui
correspond à la force active du muscle (élasticité)
Les récepteurs IAIRES fusoriaux Ia et
les récepteurs tendineux sont capables de produire des récepteurs à la fois
dynamiques et statistiques. S’il y a une augmentation brusque de la tension
musculaire (force de traction), il y a une réaction dynamique qui diminue en quelques
millisecondes (= réponse transitoire)
L’information sur
les variabilités de tension remonte de la moelle épinière au cervelet par le
faisceau spino–cérébelleux, puis elle passe par d’autres voies vers le cortex.
Ø 
Schéma
de décharge dans le fuseau neuromusculaire et l’organe tendineux :
A : Le muscle est au repos, mais
l’étirement n’est pas nul.
Les
fibres Ia déchargent tandis que les fibres Ib ne déchargent pas (seuil de
dépolarisation plus élevé) La force de traction (P1) est
insuffisante pour faire émettre des PA : les fibres Ib restent
silencieuses.
B : Le muscle est étiré au–delà de son état
de repos.
Les
fibres Ia déchargent à une fréquence plus élevée. Les fibres Ib déchargent,
mais leur fréquence de décharge est inférieure de celle des fibres Ia. Si on
cesse d’étirer le muscle, alors il y a une baisse de la fréquence des fibres Ia
et Ib.
C : Il y a une
contraction isotonique des fibres extracellulaires par stimulation des
motoneurones a.
Les
fuseaux neuromusculaires ne sont plus étirés, les fibres Ia sont silencieuses.
Les organes tendineux sont étirés du fait de la force de traction P2 (même qu’en B) Cela entraîne une réponse qui
consiste en une augmentation temporaire du taux de décharge.
Le
fuseau neuromusculaire mesure la longueur du muscle, l’organe tendineux informe
sur la tension de décharge.
D : Il y a une activation des motoneurones g qui entraîne la contraction intrafusale.
La
contraction intrafusale n’affecte pas la force de traction des fibres
extrafusales. Les fibres Ib restent silencieuses, car P1 est insuffisant
pour avoir une réponse. Mais il y a un étirement de la partie centrale du
fuseau neuromusculaire qui entraîne une réponse des fibres Ia.
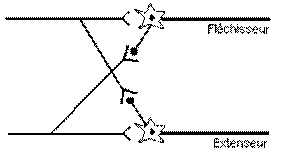
Ø Réflexe
d’étirement et d’inhibition antagoniste réciproque :
§ Flexion et extension des
membres supérieures au niveau du coude :
Les
muscles impliqués sont le biceps et le triceps.

Les
fuseaux neuromusculaires tendent à maintenir constant la longueur du muscle. Le
fonctionnement des muscles antagonistes collabore avec le fonctionnement des
muscles agonistes.
Un
poids sur l’avant bras entraîne une extension du biceps. Cela entraîne une
augmentation de l’activité du fuseau neuromusculaire du biceps qui provoque une
excitation plus intense des motoneurones du muscle fléchisseur (F) et une inhibition des motoneurones
du muscle extenseur (E)
Pendant que le
biceps (F) est étiré, le triceps (E) est, simultanément, légèrement
relâché. Ce raccourcissement passif du triceps entraîne une inhibition des
fuseaux neuromusculaires de ce muscle. Cela crée un signal Ia faible, ce qui
provoque une excitation homonyme faible via le muscle extenseur et une
inhibition réciproque des motoneurones du triceps également faible.
Cette suppression partielle ou totale de
cette inhibition est appelée désinhibition.
Arcs réflexes :
|
Arc réflexe
polysynaptique |
Ia de F |
" |
Motoneurones
de F |
" " |
Muscle
fléchisseur |
||
|
" |
Interneurones
inhibiteurs |
" |
Motoneurones
de E |
" |
Muscle
extenseur |
||
|
Ia de E |
" |
Motoneurones
de E |
" " |
Muscle
extenseur |
|||
|
" |
Interneurones inhibiteurs |
" |
Motoneurones
de F |
" |
Muscle
fléchisseur |
||
La position
relative du bras et de l’avant bras est fonction des forces exercées.
§

Généralisation :
Cet interneurone
inhibiteur reçoit aussi des entrées provenant des axones descendant des centres
cortico–spinaux. Le signal qui descend afin d’activer un ensemble de muscles
conduit immédiatement à la relaxation des antagonistes.
Par
ailleurs, d’autres voies descendantes extrapyramidales réalisent des connexions
excitatrices et inhibitrices sur les interneurones inhibiteurs.
Quand l’équilibre est déplacé vers une
inhibition plus importante sur les interneurones, l’inhibition réciproque
diminue et il y a une co–contraction. Cela permet de raidir un peu plus
l’articulation (stabilisation) pour un mouvement nécessitant beaucoup de
précision.
Ø Activité
g
et fonction de la boucle g :
§ Principe de la boucle g :
Elle
est aussi appelée innervation fusimotrice g. Elle
peut :
-
Moduler
la sensibilité du fuseau neuromusculaire et donc agir sur le déroulement d’un
réflexe myotatique,
-
Déclencher
secondairement la contraction d’un muscle initialement au repos via une
décharge Ia et une action du réflexe myotatique.

Boucle g :
fibres g " fibres Ia " fibres a
§ Activité g et
mécanisme fléchisseur–extenseur :
Fréquence
des
 afférences Ia
afférences Ia
Longueur
du muscle
fléchisseur
Le taux de
décharge des fibres motrices g est réduit sur
la courbe a
et élevé sur la courbe b. le tonus des fibres intrafusales diffère
dans les deux.
Le
taux de décharge des fibres Ia est linéaire et fonction de la longueur des
muscles.
Lorsque l’activité
des fibres g s’accroît, les
fuseaux neuromusculaires se mettent à décharger à une fréquence plus élevée (1"2) sans changement de la longueur du
muscle. L’effet de cette activité accrue des fibres Ia entraîne une excitation
plus forte des motoneurones a homonymes et une inhibition plus
forte des motoneurones a antagonistes.
La conséquence est un mouvement de
l’articulation au niveau du coude qui s’arrête dès le point 3 atteint,
c’est–à–dire que les fibres intrafusales déchargent à une fréquence égale à
celle au point 1
(avec un changement de longueur)
La
longueur des muscles peuvent peut être moduler par l’effet des efférences g. La contraction musculaire peut
être contrôlée par la boucle g et/ou par
l’activité directe des motoneurones a.
L’action directe des motoneurones a par les centres
médullaires présente un avantage : une faible latence, mais aussi un
désavantage : elle ne permet pas un contrôle délicat de la contraction
musculaire. C’est–à–dire, les fuseaux neuromusculaires impliqués peuvent être
maintenu à un taux d’étirement normal (excitation infraliminaire) ou être trop
étiré (saturation)
Ø Le
système fusimoteur maintient la sensibilité du fuseau neuromusculaire pendant
la contraction musculaire :
L’arrangement
parallèle du fuseau neuromusculaire par rapport aux fibres extrafusales pose
problème. Quand les fibres intrafusales se relâchent (quand le muscle se
raccourcit), les décharges du fuseau neuromusculaire devraient s’arrêter et
donc le fuseau neuromusculaire ne pourrait pas transmettre d’informations au
moment même où cette information est importante.
Comment fait le
SNC pour recevoir l’information
du changement de
longueur pendant la contraction ?
Travaux de Hunt et Kuffleur :
Le
SNC peut activer les motoneurones g pendant la
contraction pour maintenir une tension des fibres intrafusales du fuseau
neuromusculaire.
Hunt et Kuffleur ont étudié l’activité
des afférences de fuseau isolé avec la stimulation des fibres g du fuseau neuromusculaire et des
motoneurones a innervant les fibres
extrafusales.
Résultat :
-
Quand
il y a la stimulation des motoneurones a seuls, il y a
une pause des PA sur les afférences Ia pendant la contraction (le fuseau
neuromusculaire reste déchargé)
-
Quand
il y a la stimulation d’un motoneurone g innervant le
même fuseau neuromusculaire en même temps que le motoneurone a, il y a
l’absence de pause des PA.

Travaux de Granit :
La stimulation
électrique du cortex moteur (et d’autres centres plus élevés) entraîne la
stimulation simultanée des motoneurones a et g. C’est la co–activation a–g.
Travaux de Vallbo et Hagbarth :
Les
techniques de microneurographie permettent d’étudier l’activité des neurones
afférents dans les nerfs périphériques, de sujets humains éveillés, de
différents fuseaux pendant un mouvement volontaires.
Les motoneurones
g ont un diamètre trop petit, leur
activité électrique est impossible à identifier. L’action des neurones g doit être déduite de l’activité
des fibres Ia pendant la contraction volontaire.
L’action des
motoneurones a peut être identifiée pendant un
enregistrement électromyographique (EMG) Pendant les mouvements volontaires,
les fibres afférentes Ia augmentent souvent de fréquence de décharge, même quand
le muscle se raccourcit.
Les neurones g doivent être activé de façon
synchrone avec les neurones moteurs a. Cela confirme
que pendant une contraction volontaire, la co–activation a–g permet le maintien de l’activité
du fuseau.
Utilité de cette co–activation a–g :
On
effectue un enregistrement de la fréquence de décharge d’une fibre Ia d’un
muscle d’un doigt d’un sujet qui essaie une flexion lente et à une vitesse la
plus constante possible.
La trajectoire
du mouvement présentait des petites variations par rapport à une vitesse
constante : à certains moment, le muscle se raccourcit plus vite qu’à
d’autres.
L’activité de l’afférence Ia reflète la
vitesse de flexion :
-
Quand
la vitesse de flexion augmente transitoirement, la décharge des fibres Ia
diminue de fréquence,
-
Quand
la vitesse de flexion diminue transitoirement, la décharge des fibres Ia
augmente de fréquence.
Variation
de vitesse relative :
La fréquence de
l’afférence Ia est très sensible aux variations de la vitesse de changement de
longueur du muscle. Cette information est utilisée par le système nerveux pour
compenser les irrégularités et lisser le mouvement.
Cette information dépend de la
co–activation a–g.
Ø Réponses
des fibres ou neurones g
(moteurs) statiques et dynamiques :
Travaux de Matthews :
Il
fait des enregistrements d’activité de fibres afférentes Ia isolées avec un
étirement du muscle à vitesse contrôlée et la stimulation des axones des
motoneurones g individuels.
-
La
stimulation de certains axones g augmente
nettement la décharge d’état stable à partir de l’afférence IAIRE sur la phase
statique et sur la phase dynamique.
-
La
stimulation de certains axones g augmente
nettement la décharge durant la phase dynamique.
-
Quand
les motoneurones statiques sont activés, l’information monte dans le fuseau et
reflète essentiellement la longueur du muscle.
-
Quand
les motoneurones dynamiques sont activés, l’ensemble du fuseau devient plus
phasique, c'est–à–dire qu’il informe essentiellement sur les variations rapides
du muscle.
Ø Le
système nerveux module–t–il la contraction des fibres intrafusales
indépendamment des fibres extrafusales :
Travaux de Prochazka et Hulliger :
Au
cours d’un mouvement naturel chez le chat, il y a plus qu’un contrôle g et, dans celui–ci, il n’y a pas
qu’un lien invariant entre les activités a–g.
L’amplitude et
le type d’activité g (statique ou
dynamique) sont pré–réglés à un niveau relativement stable mais ce niveau varie
en fonction de chaque tâche spécifique et du contexte où chaque tâche est
faite.
En général, les neurones g sont régulés, activés, ajustés,
à des niveaux plus élevés lorsque la vitesse et la difficulté du mouvement
augmentent.
Exemple :
Dans
une condition imprévisible (quand le chat soulevé ou pris dans la main), il y a
une activation marquée (élevée) des neurones g.
|
Chat : |
Repos |
Assis |
Position
debout, observation |
Marche lente |
Marche rapide |
Chat soulevé
par une main |
Gratte le sol
avec les pattes arrière |
Marche sur une
poutre |
|
g dynamique : |
0 |
0 |
0 |
0 |
+ |
+ + + |
+ + + |
+ + + |
|
g statique : |
0 |
+ |
+ |
+ + |
+ + + |
+ |
+ |
+ + + |
Les
neurones g statiques
permettent un mouvement lent et prévisible.
Les neurones g dynamiques permettent un
mouvement rapide et imprévisible (mouvement brusque), lors d’un comportement
pour lequel la longueur des muscles peut changer rapidement et de façon
imprévue.
En ajustant
l’équilibre entre l’activation des neurones g statiques et
dynamiques, le système nerveux utilise le système fusimoteur pour bien accorder
et ajuster l’action des fuseaux neuromusculaires pour que le signal de sortie
fournisse l’information adéquate par rapport à la tâche spécifique à accomplir.
v Réflexe myotatique
inverse :
Ø Le
réflexe :

Les
récepteurs tendineux sont mis en jeu et réalisent des connexions inhibitrices
avec les motoneurones homonymes et des connexions excitatrices avec les
motoneurones antagonistes. Il n’y a pas de liaisons mono–synaptiques (toujours
au moins 2 synapses)
Il y a des
connexions intra–segmentaires des fibres Ib venant des organes tendineux d’un
muscle. L’étirement du triceps provoque l’utilisation de se propre contraction.
Les
connexions excitatrices entre les fibres Ib du muscle fléchisseur et les
motoneurones du muscle extenseur ne sont pas indiquées car cette voie n’existe
pas dans toutes les articulations.
Dans ce réflexe, l’augmentation de
tension (force de traction) résultant d’un étirement et/ou d’une contraction
produit l’inhibition des motoneurones homonymes par les fibres Ib. Les organes
tendineux de Golgi protègent l’organe contre toutes les charges et les
augmentations trop rapides de la tension sur les muscles et les tendons.
Ø Généralisation :
Il
y a 3 caractéristiques :
-
Les
connexions avec les motoneurones se font par des interneurones,
-
Les
afférences Ib réalisent des connexions faibles avec les muscles fléchisseurs et
se connectent fortement avec les muscles extenseurs.
-
Les
connexions des afférences Ib sont beaucoup plus étendues dans la moelle
épinière et vers les muscles somatiques que les fibres Ia.
Les actions
réflexes du système afférent des fibres Ib ne sont pas opposées à celles du système
afférent des fibres Ia, mais elles sont qualitativement différentes.

Schéma de généralisation :
Ø Phénomène
dit du « Réflexe d’Eustache » :
L’accroissement
d’étirement subi par un muscle poussé à une certaine tension provoque la chute
soudaine de la tension musculaire est attribuée à la fonction inhibitrice des organes
tendineux de Golgi. Cela permet une protection contre la surcharge.
En plus, la
baisse de la tension mécanique (force) amène une décroissance de la fréquence
des influx dans les fibres Ib, entraînant la désinhibition des motoneurones
homonymes. A ce moment–là, la tension peut à nouveau croître.
L’arc réflexe des organes tendineux de
Golgi est organisé de telle sorte qu’ils tendent à maintenir constant la
tension musculaire.
v Conclusion :
Ø Les
rétro–actions de la longueur et de la force agissent ensemble pour contrôler le
niveau du tonus musculaire :
Les muscles
agissent sur des charges pour produire à la fois des variations de longueur et
de force. Les signaux de commande centraux peuvent agir sur les rétro–actions
de la longueur et de la force (à la fois) en agissant respectivement sur les
fuseaux neuromusculaires via les motoneurones g et sur les
interneurones (rétro–action de la force)

Ø Les
interneurones inhibiteurs du groupe Ib reçoivent une entrée convergente venant
de plusieurs types de récepteurs :
L’organe
tendineux de Golgi fournit un mécanisme de rétro–action positif pour la
régulation de la tension parallèle à la rérto–action des fuseaux
neuromusculaires. Ce système tend à s’opposer aux petites variations de la
force de traction musculaire en augmentant ou en diminuant l’inhibition exercée
sur les motoneurones.
Les
interneurones inhibiteurs Ib reçoivent des entrées convergentes qui sont :
-
Des
afférences Ib,
-
Des
afférences cutanées à bas seuil,
-
Des
afférences d’articulation,
-
Des
afférences inhibitrices ou excitatrices descendantes (provenant du tronc
cérébral ou de plus haut)
Ces
connexions ont des applications fonctionnelles importantes car elles
constituent un mécanisme spinal (pour l’essentiel) pour le contrôle fin des
mouvements exploratoires tels que le toucher actif.
Quand une main entre en contact avec un
objet physique, la force musculaire sera fortement inhibée par des afférences
tendineuses et des afférences cutanées, ce qui autorise une réduction immédiate
de la force exercée pour affiner le contact.
Comme les voies
descendantes modulent aussi l’inhibition des fibres Ib, l’effet inhibiteur peut
être modulé, adapté, régulé et amène à un mouvement adéquat.
v Introduction :
expérience, résultat, récepteurs :
On
peut étudier les fonctions des centres moteurs du tronc cérébral (mésencéphale,
pont et bulbe) par section des connexions du tronc cérébral et des centres
moteurs supérieurs.
Les centres
moteurs du tronc cérébral sont responsables du contrôle des réflexes, de la
posture et de l’équilibration spatiale (3D) du corps. Ils évaluent les signaux
afférents qui leur parviennent d’un grand nombre de récepteurs situés un peu
partout dans le corps.
Ces récepteurs
sont en grande majorité :
-
Des
récepteurs des organes de l’équilibre (= organes vestibulaires de l’oreille
interne),
-
Des
récepteurs d’étirement,
-
Des
récepteurs articulaires de la musculature du cou,
-
Des
détecteurs de déformation tissulaire.
Les afférences
venant des récepteurs permettent au tronc cérébral de fournir en retour un flux
continuel d’informations motrices vers la périphérie. Les informations assurent
l’établissement et le maintien d’une posture corporelle sans intervention
volontaire.
Détection des déformations
tissulaires :
Il
existe des récepteurs dans la peau (mais pas que dans la peau) :
-
Les disques de
Merkel,
qui permettent une sensibilité tactile à la racine des poils (sensibilité des
mouvements des poils) ;
-
Les terminaisons
annexées aux poils
(= terminaisons nerveuses libres) qui s’enroulent autour des poils ;
-
Les corpuscules
de Pachini,
situés dans l’hypoderme, le périoste, à proximité des articulations, à la
surface des tendons et des aponévroses ;
-
Les corpuscules
de Meissner,
qui sont présents sur la face plantaire et la pulpe des doigts ;
-
Les corpuscules
de Krauss,
qui sont présents dans la peau, les muqueuses, les capsules articulaires,
l’enveloppe des organes internes et l’adventice des grosses artères.
Dans la peau, il
y a une sensibilité tactile mais aussi une sensibilité à la pression et aux
vibrations (fréquences de vibration des molécules de l’objet)
Dans les tissus
profonds, il y a une sensibilité d’étirement, de pression et de déformation
tissulaire.
v Relation au niveau du tronc
cérébral : organes de l’équilibration :
Ø Afférences
vers les centres moteurs du tronc cérébral :

Ø Equilibration
et organe de l’équilibre :
Il
y a 2 sortes d’équilibrations :
-
Equilibre
statique : orientation du corps en particulier de la tête par rapport au
sol (gravité)
-
Equilibre
dynamique : maintien de la position du corps en réaction à des mouvements
soudains tels que l’accélération, la décélération ou la rotation.
Le
labyrinthe est constitué :
-
D’un
lobe osseux (= cavité osseuse creusée dans la partie pétreuse du lobe temporal)
qui est divisé en 3 parties : le vestibule, les canaux semi–circulaires et
la cochlée (qui est l’organe de l’audition)
-
D’un
lobe membranaire (= réseau contenant des poches d’endolymphe) qui tapisse le
lobe osseux.
De
l’arrière vers l’avant, on trouve :
-
Les
conduits semi–circulaires,
-
Le
vestibule constitué de l’utricule et du saccule,
-
Le
conduit cochléaire (= organe de l’audition)
Les
récepteurs de l’équilibre sont les macules, situées sur la surface interne du
saccule et de l’utricule, et les crêtes ampullaires des canaux
semi–circulaires.
Toutes ces cavités sont emplies de
liquide.
Ces
éléments sont concernés par le maintien de l’équilibre et fournissent des
inforamations sur la position de la tête dans l’espace, l’accélération
ampullaire (rotation de la tête) et l’accélération linéaire.
Les accélérations positives et négatives
sont signalées également par des trains de PA (dépolarisation ou
hyperpolarisation) Les accélérations fortes, non familières, peuvent avoir des
post–effets pendant quelques secondes. Cela souligne l’inertie des mécanismes
labyrinthiques.
Chaque macula
est recouverte d’une couche gélatineuse avec des cristaux de carbonates de
calcium (= otolithes) Elle contient des cellules ciliées qui sont sous la
couche gélatineuse. Ces cellules font synapse avec le prolongement périphérique
qui forme le nerf vestibulaire.


Récepteur
sensoriel ampullaire :
Chaque
canal semi–circulaire s’élargit à l’une des extrémités pour former une ampoule
remplie d’endolymphe.
Chaque ampoule contient une crête
ampullaire avec une masse gélatineuse appelée cupule.
Le liquide passe
dans un sens ou l’autre. Un mouvement dans le sens descendant stimule les
cellules ciliées. Un mouvement dans le sens inverse inhibe les cellules
ciliées.
Disposition des
canaux semi–circulaire :
Il y a 3
canaux : un supérieur, un postérieur et un extérieur (= horizontal) qui
sont disposés perpendiculairement les uns aux autres dans les 3 plans de
l’espace. Le canal extérieur présente un angle de 30° par rapport au sol.
Sensibilité
directionnelle des cellules ciliées :


Cela concerne
les cellules ciliées maculaires et cupulaires.
La fonction des
canaux semi–circulaires horizontaux est d’apporter des informations … du
mouvement de la tête, c’est–à–dire que la vitesse et l’amplitude déterminent la
manière dont les cils sont courbés et donc les cellules ciliées stimulées.
Chaque récepteur
a un sens de sensibilité maximale. Quand les cils sont recourbés dans ce sens,
les cellules ciliées sont dépolarisées. S’ils sont recourbés dans le sens
inverse, les cellules ciliées sont hyperpolarisées.
(du nerf
8)

Quand
la tête tourne à gauche, le liquide dans les canaux est en retard sur le
mouvement de rotation à cause de son inertie. Par conséquent :
-
Le
liquide du canal gauche courbe les cils dans le sens de l’axe de
dépolarisation. Les cellules ciliées sont alors dépolarisées, libérant des neurotransmetteurs.
-
Dans
le canal droit, les cils sont courbés dans le sens inverse de l’axe de
dépolarisation. Les cellules ciliées sont alors hyperpolarisées.
Les 2
informations arrivent au cerveau (via la paire de nerf 8)
En absence de
mouvement de la tête, il existe une fréquence de décharge de base.
L’utricule et le
saccule apportent des informations sur l’accélération linéaire tandis que les
canaux semi–circulaires (récepteurs ampullaires) informent sur les
accélérations rotatives (ou angulaires) L’intégration de ces différentes
informations permet un positionnement de la tête par rapport à l’effet de
l’apesanteur.
Dans
la crête ampullaire, il existe 2 types de cellules ciliées.
Il existe, dans
la structure de la macula, quelque chose de particulier : les cellules
ciliées sont toutes dirigées vers la striola. Elles ont donc toutes le même axe
de dépolarisation (même direction mais sens différent) Elles sont donc soit
excitées, soit inhibées (+ un 3ème type qui ne répond pas ou peu)

Ø Réflexe
posturo–vestibulaire :
Il est déclenché
par des changements d’orientation dans l’espace. Il anticipe les déséquilibres
et réalise des ajustements posturaux préventifs.
Exemple :
Une chute vers
l’avant déclenche une brusque extension dans les membres antérieurs avec un
raidissement des muscles de la nuque. Cela permet d’éviter que la tête cogne le
sol.
Ce réflexe a
aussi un rôle dans la locomotion, comme dans le galop d’un cheval.
Ø Connexions
neuronales entre l’appareil vestibulaire et le SNC :
La plupart des
fibres vestibulaires se terminent dans les noyaux vestibulaires situés à
proximité à la jonction bulbo–protubérantielle (dans le tronc cérébral)
D’autres fibres vont dans le cervelet (dans les noyaux fastigiaux et dans le
lobe flocculo–nodulaire)
La
voie principale des réflexes d’équilibration, en naissance dans les nerfs
vestibulaires, part ensuite dans le noyau vestibulaire et le cervelet. Ensuite,
il y a un aller–retour intense entre le noyau vestibulaire et le cervelet.
Les influx sont transmis à la formation
réticulée du tronc cérébral ainsi qu’à la moelle épinière. Les PA à destination
médullaire permettent le contrôle de facilitation et d’inhibition de
contraction de muscles extenseurs.
Le lobe
flocculo–nodulaire du cervelet est impliqué dans la fonction
d’équilibration des canaux semi–circulaires. S’il y a la destructions de ces
lobes, les symptômes sont les mêmes que s’il y a la destruction des canaux
semi–circulaires : perte de l’équilibre qui prédomine lors d’un mouvement
brusque (l’équilibre statique est relativement conservé)
Les
signaux ascendants partent du cervelet et des noyaux vestibulaires vers la
partie haute du tronc cérébral par la bandelette longitudinale postérieure. Ils
assurent une adaptation de la position des yeux à toute rotation de la tête.
Il existe d’autres signaux ascendants
passant par cette bandelette qui vont jusqu’au cortex cérébral et se terminent
dans les aires IAIRES vestibulaires
(dans la profondeur de la scissure de Sylvius) Ils assurent la perception de
l’équilibre.
Quand les
informations sont traitées, les noyaux vestibulaires transmettent les messages
résultant au centre régissant le contrôle vestibulomoteur et spinomoteurs.

v Des neurones moteurs commandent
la position et la vitesse des yeux :
(Contrôle volontaire du regard fixe)
Ø Introduction :
Les
centres les plus élevés contrôlent le regard fixe (= gaze) et indiquent seulement un changement voulu de la position de
l’œil.
Ce signal est transformé par les
interneurones dans la formation réticulée du tronc cérébral pour devenir les
informations nécessaires de vitesse et de position pour les neurones moteurs.
Ils permettent une commande des muscles extrinsèques des yeux (globe oculaire)
Ø Activité
d’un motoneurone lors d’une saccade (= mouvement) oculaire volontaire :
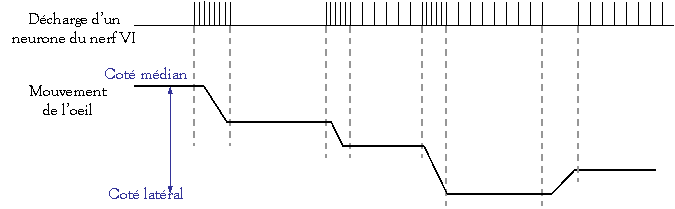
Le
motoneurone du nerf VI émet une bouffé d’activité qui précède le mouvement et
dure autant que lui.
Une augmentation du niveau tonique de la
décharge s’accompagne d’un déplacement latéral plus marquée de l’œil.
La
fréquence de décharge d’un motoneurone extra–oculaire est directement
proportionnelle à la position et à la vitesse de l’œil : tandis que la
vitesse de l’œil monte de 0 (repos) à 900°/s, la fréquence de décharge monte
rapidement démontrant une activité de pulse (= impulsion)
La montée rapide
de l’activité neuronale fait que les yeux vont aussi vite que possible et
permet d’éviter le freinage visqueux de l’orbite.
Quand la position est atteinte, l’œil
est maintenu dans cette nouvelle position par une contraction stable des
muscles extra–oculaires.
La différence de
décharge entre le niveau initial et le niveau final est appelée step.

Position de l’œil
Vitesse de l’œil
Spikes /s
(ou PA /s)
Les
saccades sont associées :
-
D’une
part avec une activité de « step » qui commande la position de l’œil,
-
D’autre
part avec une activité de « pulse » qui commande la vitesse de l’œil.
Ø Les
saccades horizontales sont générées dans la formation réticulée pontique :
Les
neurones qui réalisent la composante de pulse sont appelés les cellules de
burst (= salve) Pour les saccades horizontales, ces cellules se trouvent dans
la région para–médiane de la formation réticulée.
Elles déchargent à une fréquence élevée
juste avant et pendant une saccade ipsilatérale. Leur activité ressemble à la
composante de pulse de la décharge des neurones moteurs.
Il
y a l’existence de plusieurs types de cellules de burst :
-
Les
neurones de « long–lead
burst » qui reçoivent une entrée excitatrice des centres plus
élevés,
-
Les
neurones de burst inhibiteurs qui
sont situés plus caudalement. Ils suppriment l’activité des neurones du noyau
abducens controlatéral et suppriment aussi l’activité des neurones de burst
excitateurs
-
Les
neurones omnipause qui déchargent
continuellement sauf autour et pendant l’instant d’une saccade. La stimulation
de ces neurones pendant une saccade a pour effet d’arrêter cette saccade.
Ce sont des
neurones GABAnergiques qui inhibent les neurones de burst. Leur arrêt permet la
mise en place d’une saccade par les neurones de burst.
Une saccade nécessite
l’excitation des neurones de burst et l’inhibition des neurones omnipause.
C’est un système stable et les saccades non volontaires sont rares.
L’intégration
neuronale de la vitesse requiert le flocculus cérébelleux et les 2 noyaux du
tronc cérébral (le noyau vestibulaire médian et le noyau prépositus
hypoglossi)
On y trouve des
neurones toniques qui maintiennent un signal stable (= step) en relation avec
la position de l’œil. Ils ne possèdent aucun signal de salves saccadiques (=
burst)
Une lésion entraîne la réalisation de
saccades normales mais, après ces saccades, les yeux reviennent en position
médiane.
La composante de
la vitesse de l’œil vient essentiellement des neurones burst excitateurs qui
sont dans la formation réticulée pontique paramédiane et qui fait synapse sur
les motoneurones finaux et des interneurones du noyau abducens.

v Les réflexes vestibulaires
« stabilisent » les yeux et le corps lorsque la tête bouge
(exemple : réflexe vestibulo–oculaire) :
Ø Les
réflexes vestibulo–oculaires compensent les mouvements de la tête :
La
perception d’images stables est meilleure que la perception d’images en
mouvement. Lorsque la tête bouge, les yeux restent immobiles par rapport à la
cible via ces réflexes vestibulo–oculaires des muscles des yeux.
Le réflexe rotationnel
vestibulo–oculaire compense la rotation de la tête et reçoit une entrée
prédominante dans les canaux semi–circulaires.
Ø Le
mystagmus vestibulaire rétablit la position des yeux pendant une rotation
soutenue de la tête :
Mystaso = je m’incline
Quand les canaux
semi–circulaires détectent une rotation de la tête dans un sens les yeux
tournent lentement dans le opposé. Les yeux restent fixes par rapport à la
cible.
Hypothèse : Une rotation
soutenue dans un sens amènerait les yeux dans le bord de l’orbitre et y
seraient maintenus.
NON
" Les yeux réalisent un mouvement de mise à zéro
rapide (saccade involontaire)
Le
mystagmus est une séquence composée de mouvements (phases) lent et rapide. Le
signal vestibulaire contrôle la phase lente et les circuits du tronc cérébral
produisent la phase rapide.