
Retour au sommaire des cours |

Une version pdf est à votre disposition ici. |
Elle
consiste, avec la reproduction, l’une des propriétés caractéristiques des êtres
vivants. Si la reproduction assure la survie à long terme de l’espèce, la
nutrition assure la survie immédiate de l’individu.
Rôle
de la nutrition :
-
Renouvellement
des molécules constitutives de l’organisme en apportant les métabolites de
base,
-
Couverture
des dépenses énergétiques liées au métabolisme cellulaire, à la production de
chaleur ou aux mouvements de l’organisme.
Etapes
caractéristiques :
-
Prise
de nourriture dans le milieu = Ingestion.
-
Fragmentation
chimique et mécanique des aliments = Ingestion, absorption, distribution.
-
Rejet
hors de l’organisme des éléments non digérés et des catabolites.
v Modalité de la prise de
nourriture :
Les animaux sont
des organismes hétérotrophes : ils sont totalement tributaires des
organismes autotrophes (procaryotes et végétaux chlorophylliens)
La matière organique se trouve sous
différentes formes, avec le développement de nombreuses adaptations chez les
animaux pour prélever ces différentes formes de la matière organique présentes
dans un écosystème.
La formation de
vacuoles digestives nécessite :
-
Déformation
de la cellule permettant l’internalisation des particules phagocytées, le
déplacement intracellulaire des organites et l’émission des pseudopodes.
" Intervention de complexe d’actine, de
cytokine et d’ATP.
Les
microphages : – Suspensivores (particules en suspension dans l’eau)
– Déposivore (particules dans ou à la surface
des sédiments)
Les
macrophages : – Brouteurs : Herbivores ou carnivores
(ex : gastéropodes)
–
Prédateurs : Carnivores
–
Nécrophages : Carnivores
Ø Les
animaux microphages :
= Les animaux
consommant des particules de petite taille (inf. à 50 µm)
Les
planctophages consomment le phytoplancton (microalgues) et le zooplancton
(larves, protozoaires)
Les débris de toutes sortes (animaux et
végétaux) qui constituent le seston
représentent une biomasse d’une dizaine de fois supérieure à celle du plancton
vivant.
§ Les suspensivores :
Ils
sont aussi appelés filtreurs car ils consomment des particules qui flottent
dans le milieu aquatique en filtrant l’eau. Ils utilisent des processus qui
nécessitent :
-
La
formation d’un courant d’eau :
-
Battements
de cils ou de flagelles : ventilation des surfaces respiratoires et
déplacement (larves),
-
Mouvement
d’appendices chez les crustacés planctoniques (par exemple)
-
Filtration
des particules de manière à retenir (concentrer) les particules consommables et
à rejeter les particules indésirables.
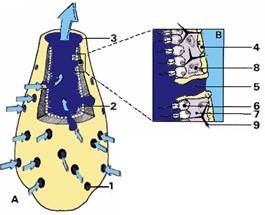
·
Paroi
et choanocytes :
Ce la concerne
les spongiaires dont le corps est traversé par un courant d’eau généré par le
battement des flagelles de choanocytes et la contraction du canal exhalant (ou
oscule)
Le
courant d’eau est optimisé par le phénomène d’entraînement visqueux.
Les particules sont phagocytées par les
choanocytes.
Les modalités
d’entrée de l’eau et de préfiltration sont variables selon les types d’éponges
(Ascon, Leucon et Sycon)

Courant :
![]()
![]()
![]()
![]()
![]()
![]()
![]()
Au raz du sol,
le flux est plus faible (résistance du fond) et est donc plus important au
niveau de l’oscule, favorisant la sortie de l’eau à ce niveau.
Tout ce qui sort dans un sens doit bien
entrer dans l’autre : cela optimise le courant d’eau.
·
Panache
tentaculaire :
Plusieurs
familles d’annélides polychètes sédentaires tubicoles possèdent une couronne de
tentacules au niveau de leur protosmium (= segment céphalique)
Leurs tentacules peuvent se dresser par
turgescence (canalicule coelomique aveugle axial) et possèdent une double
rangée de ramification (présence de pinnules)
L’épithélium des
tentacules est une association de cellules muqueuses et de plages ciliaires.
Les battements
ciliaires provoquent :
-
Un
courant de l’extérieur vers l’intérieur du panache,
-
Des
micro–courants qui entraînent les particules vers la face interne des pinnules,
puis vers le sillon du tentacule.
Le tri des
particules est fait par un système de plis situés à la base de la paroi
tentaculaire :
-
Seules
les plus petites particules atteignent le fond du sillon sont consommées,
-
Les
autres sont rejetées avec le courant exhalant.
La microphagie
par utilisation de panache est observée chez d’autres invertébrés aquatiques
mais il y a des variations possibles des mécanismes de piégeage.
Exemples :
-
Les
bryozoaires (protostomiens),
-
Les
crinoïdes (deustérostomiens/échinodernes)
·
Branchies
filtrantes :
Les mollusques : Bivalves (lamellibranches)
Gastéropodes
(rares),
Les protocordés,
Les vertébrés.
¨
Les bivalves :
Il a y 4
ordres : les protobranches, les
fillibranches, les eulamellibranches, les septibranches.
Les protobranches (ordre primitif) :
Les branchies
sont peu développées, à filaments simples. Elles n’ont qu’un rôle respiratoire
(hémostase)
Mais en général,
les bivalves (lamellibranches) se caractérisent par des branchies très
développées.
Ils sont distingués par les différents
types de branchies en fonction de la disposition des filaments : rôle dans
la nutrition (collecte des particules alimentaires)
Les fillibranches :
Les filaments de
la branchie sont solidaires les uns aux autres grâce aux cils qui se touchent.
Les eulamellibranches :
Les
filaments branchiaux sont soudés par des jonctions interfilamentaires à travers
lesquelles passent les vaisseaux sanguins : implication dans la nutrition.
Exemple : Ostréa, et les bivalves à siphon (coque, couteau,
praire, telline, etc.)
Les septibranches :
Les branchies ne
sont pas impliquées dans la nutrition. Elles sont remplacées par des cloisons
musculaires servant à pomper l’eau.
" Il n’y a que
les ordres Eulamellibranches et les Fillibranches, où les branchies sont
impliquées dans la nutrition.
Mode de collecte
des particuliers alimentaires chez les bivalves :
-
La
ciliature latérale crée un courant d’eau de la cavité hypobranchiale à travers les
branchies vers la cavité épibranchiale et vers l’extérieur.
-
Sécrétion
d’un film de mucus par des cellules glandulaires de la partie supérieure des
branchies,
-
Piégeage
et agglutination des particules,
-
Les
particules enrobées glissent vers les sillons nourriciers de chaque filament,
poussées par des courants locaux générés par des cils frontaux et
latéro–frontaux.
-
Les
particules glissent finalement le long des sillons vers les palpes labiaux et
la bouche.
Les particules
trop grosses ne peuvent pas s’insérer dans les sillons et tombent dans la
cavité hypobranchiale et sont éliminées.
Il y a un
dernier tri des particules muqueuses au niveau des palpes labiaux :
-
Les
particules digérables sont dirigées vers la bouche (courants ciliaires),
-
Le
reste sert à la une production de pseudofèces.
Chez les
protozoaires, les palpes labiaux sont très développés et constituent donc le
principal organe de capture des aliments. Ils sont par contre moins développés
chez les fillibranches et les eulamellibranches.
Ce type de
nutrition par filtration par filtration existe également chez certains
gastéropodes comme la crépidule, grâce à une branchie recouverte de mucus et
qui joue un rôle de filtre.
¨
Les procordés :
Exemple : le
branchiostoma (céphalochordés) :
Il
ressemble à un petit poisson, vit dans le sable et peut nager.
Il
possède un important pharynx brachial cilié. Les battements des cils produisent
un courant d’eau dans la bouche.
La
capture des particules alimentaire se fait dans le pharynx par un fil de mucus.
L’eau passe dans l’atrium à travers de nombreuses fentes, puis est évacuée par
l’atriopore.
Exemple d’urochordés : les ascidies :
L’adulte est
séculaire et vit fixé. La larve libre présente les différents caractères des
cordés (similaire à l’amphioxius)
¨
Les vertébrés :
La
nutrition microphagique est présente chez les larves de quelques agnathes
actuelles, comme la larve ammocète des lamproies.
La cavité pharyngienne est dilatée et
percée de 7 paires de poches branchiales qui communiquent directement avec
l’extérieur par des pores exhalants.
Mécanisme :
-
Filtration
des particules alimentaires au niveau des fentes branchiales,
-
Enrobage
par du mucus sécrété par l’épithélium pharyngien,
-
Acheminement
vers l’œsophage par des mouvements ciliaires.
La microphagie
est aussi présente chez la larves de certains amphibiens (les anoures) Il s’agit du même mécanisme que la larve
ammocète.
·
Tractus
ciliaire :
Les cils sont
disposés en rangée.
¨
Les larves
ciliées :
Il s’agit des
larves trocophores et véligères. Les cils servent à la locomotion et à la
nutrition.
¨
Les rotifères :
Ce sont des
pseudo–coelomates, de petite taille et d’eau douce, ayant un appareil rotateur
formé de couronnes ciliées impliquées dans la filtration.
¨
Les ptéropodes
et les thécosomes :
Les gastéropodes
opistobranches utilisent des expansions latérales du pied (parapodies) pour se
déplacer mais également pour collecter des particules alimentaires.
¨
Les cnidaires
scyphozoaires :
Les méduses se
nourrissent de plancton qui adhère aux 4 bras buccaux. Les cils présents sur la
face interne des bras dirigent les particules vers la bouche.
·
Appendices :
Ils sont
présents chez les crustacés (copépodes, branchiopodes et cirripodes)
¨
Les copépodes :
Leur corps est
séparé en 2 parties :
-
La
région antérieure (tête et thorax) qui porte les appendices et l’œil nauplien,
-
La
région postérieure qui est étroite et dépourvue d’appendices.
Les
copépodes représentent une grande partie du plancton marin. Ce sont des
phytophages : ils consomment des micro–algues.
Le courant d’eau
nutritif crée par le mouvement rotatif des appendices buccaux et maxillipèdes. Il
y a la présence de soies et d’épines, sur le bord interne des appendices
buccaux, qui constituent un filtre pour collecter les particules alimentaires.
¨
Les
branchiopodes :
-
Les
courants d’eau créés par les appendices thoraciques mous et aplatis sont
impliqués dans la ventilation des branchies et l’apport des particules
alimentaires par in sillon ventral passant au niveau des appendices.
-
Il
y a une agglutination des particules par une sécrétion glandulaire.
-
Puis,
ces particules sont capturées par les appendices buccaux.
¨
Les cirripèdes :
Ce sont des
formes marines fixées. Leurs appendices thoraciques biramés et allongés forment
des cirres. Ils présentent une carapace couverte de plaques calcaires (ex.
anatifes, balanes) sécrétées par l’épithélium tégumentaire.
Exemple : la
balane :
Mécanisme
mis en œuvre pour créer les courants d’eau trophiques et respiratoires :
-
Extension
du thorax (ouverture de l’opercule) et déploiement des cirres postérieures P4
et P6 (périépodes)
-
Entrée
d’eau dans la cavité palléale pour être filtrée par P1, P2 et P3.
-
Rétractation
du thorax : évacuation d’eau et enroulement des cirres qui apportent à la
bouche les particules capturées.
Les larves
d’insectes aquatiques utilisent également des soies pour capturer des
particules alimentaires.
·
Piège
muqueux :
Exemple :
le chaetoptère (infra–littoral)
-
La
nourriture particulaire est capturée par un filet muqueux sécrété par les
parapodes au niveau de la cupule.
-
Les
parapodes positionnés en éventail créent un courant d’eau.
-
La
masse muqueuse est absorbée au niveau de la bouche.
·
Organismes
filtreurs dans les écosystèmes aquatiques :
La microphagie a
une place prédominante dans le flux de matière au sein des systèmes océaniques
et lacustres :
-
Matière
organique abondante,
-
Nombreuses
espèces micorphagiques.
Les
microphages forment le plus grand groupe de consommateurs de la production IAIRE :
-
80%
de la production planctonique,
-
65%
de la matière organique dissoute, détritique ou bactérienne.
" Importance dans
l’épuration des eaux, dans l’économie comme dans la conchyliculture (bivalves)
et dans la concentration de métaux lourds et de micro–algues toxiques pour
l’homme.
NB :
Pour une moule
et pendant un an, le volume de filtration est estimé à 22 tonnes d’eau (pour
une concentration en particule environnant 5 ppm), soit
Notion de taux de filtration :
F :
Taux de filtration V :
Volume expérimentale t :
Temps expérimental C0 :
Concentration en algues au temps t0 Ct :
Concentration en algues au temps t
La
mesure est basée sur la diminution de la concentration en particules du milieu
expérimental en fonction du temps.
![]() F
= V
F
= V
t . log(C0/Ct)
Or, f(C) = – k.t
Donc
log (Ct /C0) = – k.t
log Ct – log C0 = – k.t
k = ( log C0 – log Ct ) /t =
vitesse de filtration
La filtration
est associée avec l’oxygénation. La microphagie est un processus régulé.
Les
animaux localisés dans une zone intertidale (= zone de balancement des marées)
disposent de mécanismes de compensation qui ajustent l’efficacité de la
filtration au temps pendant lequel elle est possible :
-
Réveil
rapide de l’activité de filtration,
-
Augmentation
de la vitesse de filtration.
" Influence de la taille des particules et de
la concentration en ces particules sur la filtration.
L’augmentation
de la concentration en particule entraîne l’augmentation de la quantité de
particules ingérées. Mais le phénomène est limité par la saturation du tube
digestif. Au–delà de cette limite :
-
D’abord
il y a un rejet de la nourriture en excès sous forme de pseudofécès :
sédimentation fine des milieux (vases),
-
Puis
il y a un ajustement du taux de pompage.
§ Les détritivores :
(deposit–feeders)
Ils sont aussi
qualifiés de détritivores : ils sont capables de recycler la matière
organique provenant de la fragmentation fine de la nourriture d’autres
consommateurs de débris animaux et végétaux (espèces nécrophages et microphages)
Matière
organique incorporée au sédiment (limivorie)
![]()
![]()
Matière
organique reposant à la surface ou en suspension
NB :
A la différence
de la filtration, il n’y a pas de création d’un courant d’eau.
·
Capture
par des dispositifs spécialisés :
¨
Les annélides
polychètes :
Famille
des térébellides :
-
Leur
corps est enfoncé dans les sédiments.
-
Ils
présentent des tentacules capables de balayer la surface du sable.
-
Les
détritus organiques adhèrent au mucus des tentacules.
-
Ils
sont ensuite dirigés vers la bouche par :
-
Les
mouvements ciliaires de la gouttière tentacules,
-
Et/ou
la contraction des tentacules qui possèdent une musculature propre.
¨
Les échinodermes :
Les ophiures (classes des Stelléroïdes
ou Astéroïdes) :
-
Ils
sont nécrophages, filtreurs ou détritivores.
-
De
nombreuses espèces utilisent plusieurs modes de nutrition.
-
Chez
les dépositivores :
-
Les
particules organiques sont capturées par les pieds,
-
Elles
sont ensuite dirigées vers la bouche par l’épithélium cilié entre les pieds
ambulatoires (= podia)
Remarque :
Ce sont des
organismes bio–luminescents et sont ainsi des indicateurs de métaux
lourds : la présence de métaux lourds entraîne une baisse proportionnelle
de leur bio–luminescence.
Les holothuridés (concombre de mer) :
-
Les
particules sont capturées par une couronne tentaculaire (pieds modifiés autour
de la bouche)
-
Les
tentacules peltés sont recouverts de mucus.
-
Des
mouvements de rétraction sont assurés par un afflux du liquide coelomique
associé à la contraction du muscle.
¨
Les mollusques :
Le dentale (scaphopode) :
C’est un
mollusque de forme allongée avec une coquille en forme de tube asymétrique
ouvert des 2 cotés. La tête porte des captacules préhenseurs.
-
Les
particules alimentaires sont amenées à la bouche par rétraction des captacules et
par battements ciliaires.
-
Le
pied participe également au transit grâce à son sillon médian.
Les lamellibranches protobranches
détritivores :
Exemple : la
nucule :
-
Les
palpes labiaux hypertrophiés fonctionnent comme une trompe pour fouiller le
sédiment.
-
Les
particules sont transportées par une gouttière ciliée, puis elles sont triées.
Exemple : la
telline :
Elle prélève les
dépôts particulaires par aspiration via un siphon inhalant extensible.
·
Les
limivores :
Ils sont non
sélectifs par absorption du sédiment.
¨
Les annélides :
Cela concerne
les vers fouisseurs.
Les annélides polychètes :
Exemple :
l’arénicole :
-
L’ingestion
du sédiment se fait grâce à une trompe dévaginable.
-
Un
courant d’eau pour la respiration est généré par des contractions
péristaltiques (mouvements ventilatoires)
-
L’ingestion
du sable est faite grâce à une trompe dévaginable. Toutes les 40 minutes
(environ), le vers remonte et défèque le sable (= turicule)
Les arénicoles
ont un rôle important dans la dynamique sédimentaire : 4000 tonnes par ha
et par an.
Les annélides oligochètes terricoles :
Exemple : le lombric :
Les
vers terricoles ont un rôle important dans :
-
L’aération
et l’ameublissement des sols,
-
Le
recyclage de la matière organique,
-
La
consommation des végétaux en décomposition.
= Bioturbation
D’autres vers
vivent d’avantage en surface dans la litière, le fumier ou le compost de bois
décomposé.
Les sipunculiens :
Les espèces
vermiformes portent une longue trompe rétractile avec une couronne de
tentacules courts et ciliés. Ils ont le même comportement alimentaire que
l’arénicole.
Les échinodermes :
La
classe des holothuridés et sous–classe des synaptes :
Ils sont
dépourvus de pieds ambulatoires sauf au niveau de la bouche où ils rejettent
les particules minérales.
La
classe des échinoïdes :
Il s’agit
d’oursins irréguliers dépourvus de lanterne d’Aristote.
Exemple :
Echinocardium cordatum (= oursin de sable)
-
Il
ingère le sédiment grâce à des piquants situés autour de la bouche.
-
Ce
n’est pas un limivore strict, il réalise également une sélection des détritus.
§ La panctonophagie :
·
La
baleine :
Il s’agit des
mammifères cétacés mysticètes qui se nourrissent d’animaux dont la taille
n’excède pas quelques centimètres.
La planctonophagie est proche de la
microphagie déjà décrite, si on prend en compte le rapport de taille
proie/prédateur.
Les proies sont
désignées sous de terme de krill (=
crustacés euphausiacés qui ressemblent à des crevettes mais dont les appendices
sont thoraciques)
Le
dispositif de filtration est constitué de fanons (= lames kératinisées) qui
résultent de l’allongement et la kératinisation de l’épithélium palatin.
L’animal engloutit plusieurs m3 d’eau qui sont
rejetés à travers les fanons (qui peuvent atteindre jusqu’à
·
Les
oiseaux :
Comme les flamands
roses et les pétrels, ils possèdent également des filtres de franges suspendues
à la mâchoire supérieure pour la filtration.
Þ Phénomène de convergence évolutive.
·
Les
poissons :
Cela
concerne les harengs, sardines, anchois, requins pèlerins.
-
Le
plancton est filtré par de longues épines portées par les arcs branchiaux
(branchicténies)
-
Ouvrant
largement son immense gueule, à une vitesse moyenne de
-
L’eau
est évacuée au travers des branchies.
Ø Les
animaux se nourrissant de liquide :
L’alimentation à
partir de liquide requiert le plus souvent des structures particulières
permettant le percement ou l’incision et impliquera des mécanismes assurant la
succion et/ou le léchage.
§ Percement et succion :
·
Espèces
se nourrissant de sève :
Ce sont les
insectes hétéroptères (= hémiptères) et les homoptères (puceron, phylloxera,
cochenille, cigale, etc.) Ils présentent un appareil buccal appelé le rostre.
NB :
Chez ces
insectes, si l’appareil buccal est formé par un ensemble de pièces buccales qui
suit un plan d’organisation général, il peut présenter de profondes
transformations en fonction du régime alimentaire.
L’appareil
buccal est constitué :
-
Du
labre (= lèvre supérieure = repli chitineux),
-
D’une
paire de mandibules (= appendices) abrégé : Md,
-
D’une
paire de maxilles masticatrices abrégé : Mx,
-
Du
labium (= lèvre inférieure possédant souvent une paire de pièces extérieures
appelées palpes labiaux)
Le rostre est un labre très développé,
formant un étui qui entoure les mandibules et les maxilles transformées en
longs stylets dont le mouvement assure le percement.
Les
stylets maxillaires sont creusés de 2 sillons qui, ensembles, forment des
canalicules :
-
Canalicule
dorsal : canal alimentaire assurant la succion,
-
Canalicule
ventral : canal salivaire.
·
Les
nématodes phyto–parasites :
Ils
utilisent leurs stylets creux présents dans la capsule buccale :
-
Ejection
pour percer l’épiderme des racines,
-
Pompage
des fluides par le pharynx.
Dans
le cas des espèces qui infectent les feuilles :
-
Insertion
du stylet au niveau des stomates
" Pas de dommages au niveau de l’épiderme.
·
Les
animaux hématophages :
Exemple : les annélides
hirudinées (achètes) : les sangsues :
Les ¾ des
espèces sont hématophages (les autres sont prédateurs)
-
Perforation
du tégument à l’aide d’une trompe dévaginable ou à l’aide de mâchoires
dentelées.
-
Glandes
salivaires sécrètent un anticoagulant proche de l’héparine (inhibition des
facteurs responsables de l’activation prothrombaire = thrombine)
-
Le
sang est aspiré par le pharynx.
¨
Les arthropodes :
Les
crustacés :
Cela concerne
quelques isopodes et copépodes qui s’approvisionnent sur des poissons et
crustacés.
Les
chelicérates arachniques (acariens) :
Exemple :
la tique.
-
Ancrage
par le rostre (chélicères + pédipalpe + éperon)
-
Succion
par le mouvement musculaire du pharynx
Après la
fécondation, la femelle monte à l’extrémité d’une branche de buisson et se
laisse tomber sur un mammifère lorsqu’il passe. Elle est attirée par les traces
d’acide butyrique exhalé par la peau du mammifère.
Les tiques sont
vecteurs de maladies (maladie de Lyme) dont l’agent est une bactérie de la
famille des spyrochètes (= maladie articulaire et neurologique)
¨
Les insectes :
Exemples : le moustique
femelle (diptère), la puce (siphonoptère), le pou (anoploure), la punaise de
lit (hémiptère ou hétéroptère)
Le
moustique :
-
Le
mâle se nourrit de sucs végétaux, de lipides organiques et de substrats en
décomposition.
-
La
femelle se nourrit de sang.
-
Elle
présente un labre allongé formant une trompe renfermant les aiguillons
perforants.
-
Les
mandibules + les maxilles + hypopharynx forme l’aiguillon chitineux, renfermant
lui–même un canal le long duquel s’écoule la salive.
-
Le
labre et l’hypopharynx s’insèrent dans le trou pour permettre le pompage.
§ Incision et léchage :
C’est
le cas de certaines mouches :
-
Les
mandibules sont tranchantes pour l’incision.
-
Le
labium est spongieux pour le léchage.
·
Les
chordés :
Cela
concerne les agnathes (lamproie et mixine)
La bouche est
équipée de râpes qui provoquent des blessures et se nourrissent du sang qui
jaillit de poissons malades et qui s’attaquent aux poissons malades affaiblis.
·
Les
mammifères :
Cela
concerne les chauves–souris vampires.
La blessure est
provoquée par une morsure. La salive contient des anticoagulants et des
analgésiques.
§ Butinage :
·
Les
lépidoptères :
(Papillons)
L’appareil
buccal de type lécheur–suceur :
-
Le
labre et les mandibules sont peu développés.
-
Les
maxilles forment une longue trompe enroulée qui permet l’aspiration du nectar.
-
Le
labium est peu développé.
·
Les
hyménoptères :
(Abeilles)
L’appareil
buccal de type lécheur–labial :
-
Le
labre est réduit.
-
Les
mandibules permettent le malaxage de la cire et la récolte du pollen.
-
Les
maxilles sont allongées.
-
Le
labium est allongé, long, hérissé de poils et creusé par un canal médian. Il
sert à pomper le nectar des fleurs.
§ Autres formes :
·
Le
cas des chélicérates :
Ils
ne possèdent pas d’appendices pour la mastication. Ils se nourrissent de
liquide bien qu’ils soient souvent prédateurs.
-
Les
chélicères (= homologues des antennes des crustacés et des insectes) percent la
proie.
-
L’animal
injecte des sucs digestifs.
-
Les
tissus de la proie sont liquéfiés puis aspirés.
·
Les trématodes
(= douves) :
L’orifice buccal
s’ouvre au centre d’une ventouse. L’aspiration se fait par le pharynx
musculeux.
·
Nourriture
liquide des jeunes :
¨
Les mammifères :
Le lait est
sécrété par les glandes mammaires.
¨
Les oiseaux
(columbidés) :
(Pigeons,
tourterelle)
Le
« lait » est sécrété par le jabot. Cette sécrétion est sous le
contrôle de la prolactine comme chez les mammifères.
Le lait assure la nourriture riche non
dépendante d’un approvisionnement en nourriture irrégulier ou intermittent.
·
Mécanisme
d’osmotrophie :
Il s’agit d’une
absorption cutanée de substances dissoutes, ayant pour conséquence un appareil
digestif régressé.
¨
Les cestodes :
Exemple :
le taenia : C’est un plathelminthe parasite de l’intestin–grêle de
vertébrés.
¨
Les crustacés :
Il
s’agit de cirripèdes parasites d’autres crustacés.
Exemple :
la sacculine du crabe effectue un prélèvement métabolique important au
point d’interdire toutes croissance et reproduction à l’hôte.
Ø Les
animaux macrophages :
Il s’agit des
animaux dont la nourriture constitue une masse importante comparée à leur masse
propre. On retrouve des herbivores (phytophage), des carnivores et des
omnivores.
§ Les brouteurs :
Ce sont
généralement des herbivores (ou phytophages) Il existe cependant des brouteurs
carnivores qui se nourrissent d’animaux fixés comme les spongiaires, les
ascidies ou les hydraires (exemple : les nudibranches)
·
Les
mollusques :
Ils utilisent
une radula pour râper et triturer les plantes vasculaires ou les algues dont
ils se nourrissent. La radula est un ruban associé à des dents kératinisées.
¨
Les gastéropodes
pulmonés :
La cavité
palléale s’est transformée en poumon. On retrouve des gastéropodes terrestres
(escargot, limace) ou d’eau douce (limnées, planorbes, etc.)
¨
Les gastéropodes
opistobranches :
Leur
branchie est située en arrière du cœur. On retrouve les aplysies ou les
nudibranches (= limace de mer)
Les nudibranches sont de coquille et de
parapode. Elles se nourrissent d’éponges et de cnidaires. Ils sont capables de
stocker les nématocystes des cnidaires dont elles se nourrissent.
¨
Les
polyplacophores :
Exemple : les
chitons :
Ils broutent les
algues. Toutefois, certains sont prédateurs ou détritivores.
·
Les
arthropodes :
-
Les
crustacés consomment souvent des débris végétaux (détritivores)
-
Les
insectes présentent des espèces herbivores (sauterelles, criquets) et des
espèces xylophages (termites)
Exemple : le
criquet :
Il présente un
appareil buccal de type broyeur (le plus primitif chez les insectes)
Cet appareil est constitué :
-
D’un
labre = simple repli chitineux,
-
D’une
paire de mandibules (Md) = pièces masticatrices dentées chitineuses,
-
D’une
paire de maxilles (Mx) = pièces biramées et des palpes maxillaires sensoriels,
-
D’un
labium = 2 pièces soudées portant une paire de palpes labiaux sensoriels,
Pour une même
espèces de criquet, il existe des formes solitaires et d’autres grégaires. La 1ère
forme possède des ailes plus longues et une pigmentation plus prononcée. La
forme grégaire provoque des ravages.
·
Les
équinodermes :
¨
Les équinidés :
Il
y a utilisation de la lanterne d’Arsitote chez l’oursin. Elle est constituée de
5 mâchoires articulées portant chacune 1 dent minéralisée. Les mâchoires sont
actionnées par des muscles puissants assurant :
-
La
rétractation et la protraction de la lanterne,
-
L’écartement
et l’adduction des dents.
Ils mangent des
algues et des animaux.
·
Les
vertébrés :
¨
Les oiseaux :
Ils présentent
un bec corné, de forme conique, court et robuste chez granivores.
¨
Les reptiles :
Quelques uns
sont végétariens : tortues terrestres, iguanidés. Ils possèdent des
mâchoires garnies de dents.
Þ La dentition
reflète la nature du régime alimentaire :
Dents labiales : incisives préhensives,
Dents jugales : prémolaires et molaires
masticatrices.
¨
Les mammifères :
Exemple d’herbivore rongeur : le lapin :
-
Il
possède des incisives à croissance continue taillées en fuseau,
-
Il
y a absence de canines,
-
Il
possède 3 prémolaires et 3 molaires à croissance continue.
Exemple d’herbivore ruminant : le boeuf :
-
Il
possède des dents à croissance limitée.
§ Les prédateurs :
Ce sont des
animaux zoophages. Leurs proies sont mobiles, il y la nécessité de stratégies
de capture (poursuite, pièges) et d’outils de capture.
·
La
poursuite :
Les prédateurs requièrent :
-
Des organes des
sens très développés pour la vision, l’odorat (signaux
chimiques), l’audition, la
pression :
Les requins sont
sensibles aux différences de pression, et aussi à la faible activité électrique
générée par l’activité musculaire des proies. Il s’agit d’électroréception liée
à la présence d’ampoules distribuées sur la tête.
-
Un système de
locomotion efficace :
-
La
chasse à la course, nage, vol,
-
La
chasse à l’affût (accélération)
-
Des organes
préhenseurs spéciaux :
-
Des
« bras pêcheurs » des céphalopodes comme la seiche, munis de
ventouses,
-
Des
pinces des crustacés (péreispodes antérieurs),
-
Des
griffes (mammifères et oiseaux),
-
Des
chélicères et pédipalpes des chélicérates,
-
Une
langue préhensile des batraciens et reptiles (grenouilles, crapauds, caméléons)
-
Des organes de
mastication–dilacération :
-
Un
bec de perroquet inversé des céphalopodes (composé de chitine et de
scléroprotéine, avec la présence d’une radula sans rôle essentiel),
-
Une
denture :
-
Des
requins (poissons cartilagineux de la classe des chondrichtyens),
-
Des
poissons téléostéens,
-
Des
reptiles (crocodiles),
-
Des
mammifères carnivores (félins) :
-
Développement
de canines (= crocs),
-
Prémolaires
pointues et tranchantes,
-
Molaires
atrophiées (une seule par demi mâchoire)
-
Absence
de dents broyeuses,
-
La
mâchoire inférieure n’effectue que des mouvements verticaux.
La
dent est une différenciation épithéliale.
Chez les
vertébrées non mammaliens (sauf les serpents venimeux), les dents ne sont pas
différenciées : maintien de la proie, déchiquetage et ingurgitation.
-
Des glandes à
venin (chez
certaines espèces) viennent s’ajouter à
cet arsenal :
-
Aiguillon
des hyménoptères,
-
Aiguillon
(dard) des scorpions,
-
Glandes
venimeuses des pédipalpes des pseudoscorpions,
-
Appareil
venimeux associé aux chélicères chez les araignées,
-
Organes
venimeux des cônes (gastéropodes),
-
Glande
salivaire des céphalopodes,
-
Appareil
venimeux des serpents.
La
production de substances toxiques sert le plus souvent à paralyser et tuer les
proies. Elle constitue également un moyen très efficace de défense et de protection
contre d’éventuels prédateurs (exemples : vives, rascasses, raies)
Certains poissons produisent au niveau
de leur peau des glycostéroïdes et des peptides neurotoxiques.
Egalement,
chez les amphibiens anoures, la peau sécrète des alcaloïdes, des stéroïdes et
des peptides qui souvent présentent des structures homologues des hormones du
cerveau et gastro–intestinales des mammifères.
Elles ont des effets neurotoxiques,
hémolytiques, cytotoxiques et présentent également une activité myotropique.
Exemple : la caeruléine
(Syrrhophus caeruleus) :
C’est un
homologue à la cholécystokinine des vertébrés qui induit la sécrétion des
enzymes pancréatiques et de la bile.
Exemple : les
kinines :
Elles provoquent
une vasodilatation et des oedèmes.
¨
Les hyménoptères
:
L’appareil
venimeux est présent chez les femelles (guêpes, abeilles, fourmis, etc.) Il
constitue les derniers anneaux de l’abdomen et est formé :
-
De
2 glandes acides,
-
D’une
glande basique,
-
D’un
appareil inoculateur = aiguillon.
Le
venin des hyménoptères contient :
-
Des
protéines neurotoxiques, hémolytiques et allergènes :
-
La
mellitine chez l’abeille,
-
Des
kinines hémolytiques et mastoparanes chez les vaspidés.
-
Des
enzymes cytotoxiques et hémolytiques :
-
Phospholipases,
-
Phosphatases,
-
Hyaluronidase
(dégrade l’acide hyaluronique = mucopolysaccharide acide, constituant du TC,
lame basale)
-
Estérases.
-
Des
amines inflammatoires :
-
Sérotonine,
-
Histamine,
-
Dopamine,
-
Noradrénaline,
-
Adrénaline.
Les
peptides bioactifs :
-
La
mellitine interagit avec les phospholipides membranaires et a un effet
hémolytique,
-
L’apamine
est un neurotoxique,
-
Les
mastroparanes ont un effet sur la dégranulation des mastocytes.
L’histamine est
un puissant vasodilatateur et provoque des réactions d’inflammatoires.
Chez les
hyménoptères sociaux, l’aiguillon joue un rôle défensif. Il disparaît dans
plusieurs familles d’abeilles et de fourmis.
¨
Les chélicérates
(scorpions, pseudoscorpions, araignées) :
Exemple : le
scorpion :
Il capture sa
proie avec les pédipalpes, se courbe et injecte son venin à l’aide de son
crochet situé dans le telson. Le crochet est un dard venimeux relié à une
ampoule à venin qui renferme 2 glandes. L’inoculation résulte de la contraction
de la tunique musculaire de la glande.
Composition
du venin :
-
Des
enzymes :
-
Protéases,
-
Hyaluronidase,
-
Petites
protéines basiques (neurotoxines)
Il a un rôle de
défense contre les prédateurs et de tuer les proies (insectes et autres
arthropodes)
-
Chez
les mammifères, les toxines interagissent avec les canaux sodiques des
membranes de cellules excitables (retard d’activation de ces canaux)
-
Chez
les insectes, il y a un blocage des PA entraînant une paralysie.
-
Le
venin des pseudoscorpions n’est actif que sur de petits arthropodes.
Exemple : les
araignées :
La glande à
venin débouche au niveau des chélicères. L’expulsion des produits est reliées à
la contraction de l’enveloppe musculaire de la glande.
Composition
du venin :
-
Des
amines biogènes,
-
Des
enzymes,
-
Des
neurotoxines.
Seules
quelques espèces sont dangereuses pour l’homme comme :
-
Atrax
robustus (dans la région de Sydney),
-
Les
mygales (Europe, Asie, Afrique, Amérique),
-
Veuves
noires (Théridiidae)
Mode
d’action du venin :
-
Libération
massive d’acétylcholine,
-
Puis,
un blocage pré–synaptique de la libération d’acétylcholine qui entraîne un
blocage de la transmission neuromusculaire.
NB :
Bien que
contenant des enzymes, le venin ne participe pas (ou peu) à la digestion
extracorporelle. La digestion est assurée par les sécrétions du mésentéron.
¨
Les mollusques :
-
Les
toxines (non peptidiques) des glandes salivaires des céphalopodes,
-
Les
conotoxines peptidiques du cône (gastropodes prosobranches)
Ils chassent les
poissons, mollusques et polychètes. Ils possèdent une dent creuse à l’extérieur
d’une pompe.
Mode
d’action des conotoxines :
-
Inhibition
de la libération de l’acétylcholine : w_conotoxine,
-
Effet
post–synaptique sur les récepteurs de l’acétylcholine : a_conotoxine,
-
Blocage
des canaux Ca2+ qui entraîne la réduction de
PA : m_conotoxine.
¨
Les cnidaires :
Le poisson des
nématocystes de certains est extrêmement toxique.
Composition
du venin :
-
Histamine,
sérotonine, prostaglandines " Douleurs et inflammations.
-
Peptides
et protéines à activité cardio–, neuro– ou myotoxique et hémolytique.
¨
Les vertébrés :
les serpents (Ophidiens) :
La mâchoire :
L’organisation
de la mandibule permet la déglutition de proies volumineuses. Certaines lignées
ont développé un appareil venimeux par différenciation de certaines glandes
buccales (salivaires)
-
Les couleuvres sécrètent une
salive peu venimeuse. Chez les couleuvres de Montpellier, le venin s’écoule le
long d’un sillon creusé dans une ou 2 dents maxillaires postérieures ; la
proie est envenimée au cours de la déglutition.
-
Les cobras
(najas), les mambas et les serpents marins possèdent des dents transformées
en crochets avec un sillon. Le venin est injecté par compression de la glande
pendant la morsure.
-
Les vipéridés et
les crotalidés (serpents à sonnette) présentent un canal avec un canal de
très grande taille, mobiles au moment de la morsure.
Composition
du venin :
-
Des
enzymes à activité nécrosante et perturbation de la coagulation sanguine,
-
Des
toxines agissant au niveau de la transmission synaptique :
-
Toxines
curarisantes qui
agit sur la liaison de l’acétylcholine au récepteur nicotinique :
-
Blocage
de la transmission musculaire,
-
Mort
par arrêt respiratoire (cobras et serpents marins)
-
Toxines b1 qui bloquent la
libération d’acétylcholine :
-
Homologue
de la phospholipase A2 (basique) qui a une action myonécrotique (mambas et
serpents des bananiers)
-
Fasciculines qui inhibent
l’acétylcholine estérase :
-
Action
en synergie avec la toxine b.
-
Cardiotoxines qui sont des
polypeptides basiques qui agissent sur le muscle cardiaque.
-
Myotoxines qui nécrosent
la peau, le TC et les muscles squelettiques.
Chez les
élapsidés (cobras, mambas), il y a la présence de 3 catégories de toxines qui
ont une action foudroyante.
·
Le
piège :
Cela
concerne surtout les espèces prédatrices peu mobiles, sédentaires ou fixées.
Elles développent différentes stratégies :
-
Le
mimétisme chez les chasseurs à l’affût,
-
Les
organes pièges et pièges construits.
¨
Les
cnidaires :
Ils
possèdent une couronne de tentacules autour de la bouche (polypes), à la
périphérie de l’ombrelle (méduses) Les tentacules sont armés de cellules
urticantes ou cnidoblastes prolongés d’un cnidocil très sensible et possédant
une vacuole (cnidocystes)
" Paralysie de la
proie
" Rétraction et
enroulement des tentacules déclenchés par des molécules libérées par les
proies.
Exemple : Le
glutathion :
= a_glutamyl_cystéine_glycine
qui est synthétisée par une voie enzymatique chez les hydres, les physalies et
quelques anémones.
Autres exemples : Les acides
aminés :
Comme la
proline, la leucine, la valine, le glutamate (anthozoaires)
¨
Les araignées :
Elles élaborent
des pièges construits (élaboration d’outils)
Exemple : La toile des
araignées (quelques familles seulement) :
-
Sécrétion de
soie :
La
scléroprotéine est produite par les glandes séricigènes situées dans l’abdomen
et débouchant au niveau d’appendices rudimentaires : les filières.
-
La soie des
pièges :
Cette soie plus
gluante est produite par les glandes flagelliformes débouchant au niveau des
filières latéropostérieurs.
Exemple : l’épeire :
-
Elle
présente une construction stéréotypée avec un cadre, des fils rayonnants,
une spire centrifuge, puis une spire centripète faite de soie gluante.
-
L’araignée
reste en relation avec la toile par un fil avertisseur,
-
Une
proie empêtrée est ligotée, paralysée puis digérée.
NB :
La soie sert
également à la construction de nids, comme moyen de locomotion et de dispersion
par le vent.
Ø Autres
formes de nutrition :
§ Association
symbiotique :
Exemple :
-
Association
symbiotique de cyanobactéries avec des éponges.
-
Association
symbiotique de chlorophycées (zoochlorelles) ou dinophycées (zooanthelles) avec
des ciliés, des éponges, des cnidaires, des polychètes et mollusques.
" Les algues
fournissent des métabolites à l’animal essentiellement sous forme d’oses
(glucose, maltose), de glycérol et d’acides aminés (photosynthèse)
" Les animaux
apportent du CO2 et du NH3 qui proviennent
du catabolisme.
-
Association
symbiotique de bactéries avec des riftia (= vers herbicoles sans bouche, ni
appareil digestif)
" Les bactéries
sont hébergées au niveau d’un organe volumineux : le trophosome qui occupe la majeure partie du cœlome.
" Les bactéries
symbiotiques oxydent le SH2 (avec du H2O et de l’O2) pour former de
l’ATP.
" Par réduction
du CO2, les bactéries du trophosome
synthétisent les composés organiques qui sont transférées vers l’animal.
Chez certains
mollusques abyssaux gastéropodes et bivalves, les bactéries sont hébergées dans
les branchies ou le manteau.
v Les systèmes
digestifs :
A l’exception
évidemment des diploblastes (protozoaires, spongiaires, cnidaires) et des triploblastes
acœlomates (platelminthes), les triploblastes pseudo–cœlomates (némathelminthes)
et cœlomates ont développé un tube digestif.
Ce
tube digestif plus ou moins différencié permet le transit, la digestion
mécanique et la digestion chimique séquentielle des aliments.
Ce tube digestif possède 2 orifices pour
l’ingestion et l’élimination : la bouche et l’anus.
"
Protostomiens et deustérostomiens.
Le tube digestif
est subdivisé en tube digestif antérieur, moyen et postérieur.
-
Les
tubes antérieurs et postérieurs sont constitués d’ectoderme.
-
Le
tube moyen est constitué d’endoderme. C’est la principale région de la
digestion et de l’absorption.
Chaque région
présente des différenciations morphologiques.
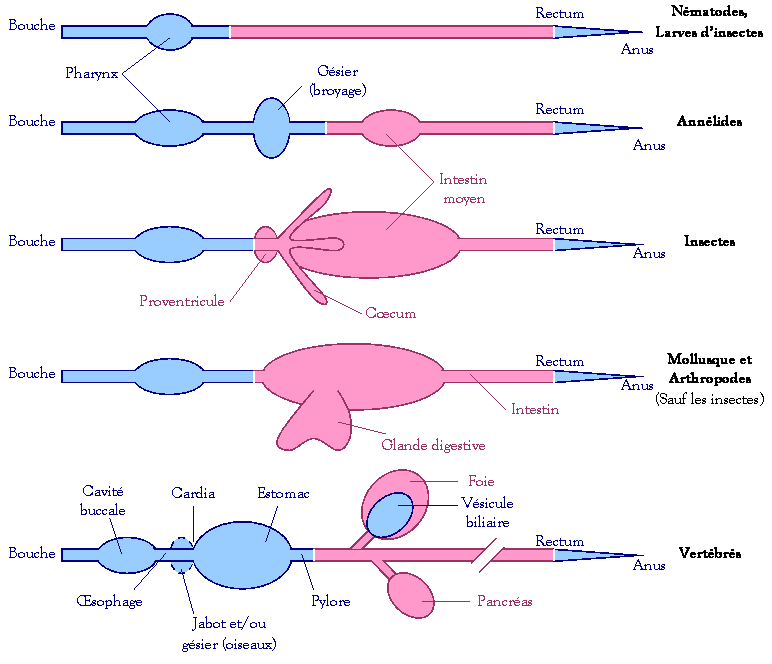
Ø Le
tube digestif antérieur (= stomodéum) :
Il contient la
cavité buccale, le pharynx, l’œsophage, le gésier et l’estomac. Il assure la
prise alimentaire : fonction souvent exercée par les organes indépendants
de l’appareil digestif.
-
Les
organes de capture d’origine buccale ou pharyngienne se trouvent le plus
souvent chez les triploblastiques primitifs (comme la trompe dévaginable
des nématodes et des annélides)
-
Pour
les phylums les plus évolués, le complexe buccal ou stomodéal participe à la
fragmentation des aliments, au stockage voire à la prédigestion enzymatique.
§ La cavité buccale :
·
Les arthropodes
:
La cavité
buccale est le lieu de transit alimentaire vers le pharynx et le lieu de
sécrétion salivaire par des glandes (transformées en glandes sérigères chez les
lépidoptères)
·
Les mollusques
:
Chez
les espèces non microphages ou suspensivores, on y trouve :
-
Des
mâchoires pour le déchiquetage,
-
Une
radula pour le broutage,
-
La
glande salivaire pour la sécrétion de mucus (mucine = glucoprotéine
enzymatique)
·
Les vertébrés
:
On
y trouve :
-
Une
langue pour le brassage des aliments,
-
Des
mâchoires avec des dents ou des odontoïdes (chez la lamproie, par
exemple) :
-
Les
dents ont une origine dermo–épidermique,
-
Les
odontoïdes sont une production épidermique kératinisée qui consiste :
-
En
des « dents » chez la lamproie,
-
En
un bec corné chez les têtards d’amphibiens.
La cavité
buccale a un rôle depuis la préhension jusqu’à la mastication chez les
mammifères.
§ Le phaarynx :
Chez les
vertébrés, le pharynx est le carrefour des voies respiratoires et des voies
digestives.
§ L’œsophage :
Il est constitué
de tissu musculeux dont le rôle est d’assurer la propulsion du bol alimentaire
par des contractions péristaltiques de la paroi (= contractions de fibres
annulaires et longitudinales)
Il
peut se dilater en jabot qui est le lieu de stockage des aliments. On le
trouve :
-
Chez
les annélides hinidinées (sangsue), où il y a un ramollissement des
aliments ;
-
Chez
les insectes herbivores, où il y a une prédigestion liée à l’action des enzymes
salivaires ;
-
Chez
de nombreux mollusques herbivores ;
-
Chez
les oiseaux, qui sert à la fermentation ou à la prédigestion des aliments qui
sont régurgités pour nourrir les jeunes au nid.
§ Le gésier :
Il
présente une paroi musculeuse qui permet le broyage des aliments pour les animaux
sans mâchoire ou appendices masticateurs.
-
Chez
les annélides (certains oligochètes ou polychètes) ;
-
Chez
les insectes qui présentent un gésier (ou proventicule) possédant un revêtement
interne cuticulaire, des épines et des crêtes ou des dents (espèces
macrophages) ;
-
Chez
des mollusques qui présentent un gésier masticateur comme chez l’aplysie
(gastéropode opistobranche) ;
-
Chez
les vertébrés (oiseaux), où le gésier constitue la partie postérieure de
l’estomac. Il contient du sable et des cailloux.
§ L’estomac :
Son
appellation est impropre chez les invertébrés.
·
Les
crustacés :
Il
y a la présence de l’appareil de tamissage chez les microphages (copépodes et
branciopodes) et la présence un moulin gastrique chez les malacostracés
décapodes (langouste, crabe, écrevisse) Ce moulin forme 2 chambres :
-
Une
chambre cardiaque (antérieure),
-
Une
chambre pylorique (postérieure)
-
La
chambre cardiaque permet un broyage secondaire des aliments, le broyage
primaire ayant été effectuée par les pièces buccales.
-
La
chambre pylorique permet un tri des particules alimentaires :
-
Les
particules les plus fines sont aspirées dans les tubules de la glande digestive
qui est en communication avec la chambre pylorique ;
-
Les
particules moyennes sont broyées par les rangées de soies ou
entraînées dans l’intestin ;
-
Les
grosses particules sont broyées par les dents latérales et dorsales.
·
Les
mollusques :
¨
Les gastéropodes
macrophages :
Ils présentent
une radula, un jabot (ou gésier) et un estomac où s’effectue une digestion par
des enzymes provenant de la glande digestive.
¨
Les céphalopodes
:
L’estomac est
l’organe de broyage lié aux contractions violentes de la paroi. La digestion
s’effectue par le déversement d’enzymes par la glande digestive.
¨
Les mollusques microphages
:
L’estomac
présente :
-
Un
bouclier gastrique chitineux (qui est abrasif),
-
Un
stylet cristallin (sécrété par le sac cristallin) qui est formé d’une matrice
protéique, de glycosidases et de lipases.
-
Le
sac du stylet cristallin dont les cellules ciliées entraînent la rotation du
stylet.
L’estomac
effectue une attaque mécanique et enzymatique du cordon muqueux (contenant les
particules alimentaires) provenant de l’œsophage.
·
Les
vertébrés :
L’estomac
est absent chez :
-
Les
cyclostomes (agnathes, myxine, lamproie),
-
Les
chimères (classe des chondrichtyens/sous–classe des holocéphales/),
-
Les
dipneustes (classe des ostéichtyens/sous–classe des sarcoptérygiens/),
-
Et
de nombreux téléostéens.
L’estomac est
caractérisé par un épithélium muqueux qui sécrète le mucus dans la lumière.
L’épithélium s’invagine et forme des cryptes glandulaires tubuleuses. La nature
des produits de sécrétion dépend de la région de l’estomac.
-
Cardia : Cellules à sécrétion de muqueuse (chez
certains mammifères),
-
Fundus : Cellules principales qui sécrètent des
pepsinogènes,
Cellules
bordantes qui sécrètent des HCl,
-
Pylore : Cellules sécrétrices de mucus et d’un peu
d’enzymes.
Rôle de l’estomac :
-
Le
stockage,
-
Le
brassage lié à la contraction de la musculature,
-
La
digestion avec du HCl (pH 1) qui favorise la formation de pepsine (=
endonucléase)
La
pepsine hydrolyse les liaisons peptidiques entre :
-
Les
acides aminés aromatiques (tyrosine et phénylalanine),
-
Les
acides aminés dicarboxyliques (aspartate et glutamate)
Il y a également la production de
chymosine ou de présure (qui coupe la caséine du lait) chez les jeunes
mammifères.
¨
Les oiseaux :
L’estomac
antérieur est un ventricule succenturié (glandulaire sécréteur)
L’estomac
postérieur est le gésier qui présente une contraction rythmique de la paroi musculaire
et une cuticule protectrice (constituée de scléroprotéine : la koiline)
Il sert de barrage aux poils, plumes et
os. Chez les rapaces diurnes et nocturnes, il y a l’expulsion par sous forme de
pelote de réjection.
¨
Les ruminants :
Chez la plupart
des herbivores, il y a un estomac uniloculaire sauf dans le sous–ordre des
ruminants (moutons, cerfs, chèvres, chameaux, antilopes, girafes, bœufs, …)
Il
y a 4 chambres : la panse (rumen), le bonnet (réticulum), le feuillet
(omasum), la caillette (abomasum)
-
Les
3 premières chambres correspondent à une dilatation de l’œsophage.
-
La
caillette est l’homologue du véritable estomac.
-
La
panse est une chambre de fermentation (jusqu’à 250L chez le bœuf adulte) qui
héberge des bactéries (Ruminobacter
et Ruminococcus) et des ciliés
entodiniomorphes anaérobies, ainsi que des champignons microscopiques.
" Les cellules ciliées + les bactéries =
-
Les
bactéries possèdent de la cellulase et sont donc capable de dégrader la
cellulose et la lignine.
-
Les
ciliés sont également capables de dégrader les polymères végétaux, mais leur
rôle essentiel est basé sur leur activité prédatrice vis–à–vis des bactéries.
Ils contribuent au maintien de la population bactérienne dans une phase
exponentielle.
Les glandes
salivaires constituent donc, chez les ruminants, un organe impliqué dans
l’excrétion azotée.
Périodiquement,
une partie du contenu de la panse est régurgitée puis mastiqué, ce qui favorise
le broyage des fibres des fibres végétales non dégradées.
Une partie de la panse passe également
dans le bonnet et le feuillet où s’effectue une réabsorption d’eau.
Dans la
caillette (véritable estomac), les ciliés et les bactéries sont prédigérés par
le suc gastrique. Donc les ciliés et les bactéries constituent le régime
alimentaire des ruminants (source d’acides aminés essentiels, de vitamines,
etc.)
NB :
Chez les autres
herbivores non ruminants (chevaux, rhinocéros, rongeurs, etc.), la dégradation
de la cellulose est effectuée par des bactéries et des ciliés hébergés dans de
volumineux cœca intestinaux.
Ø Le
tube digestif moyen :
Si le tube
digestif antérieur est le lieu où s’effectue essentiellement un traitement
mécanique de la nourriture (en particulier chez les invertébrés où l’estomac
n’a pas d’activité enzymatique), le tube digestif moyen est le lieu où
s’effectue la digestion enzymatique et l’absorption des métabolites qui en
résultent.
Cette
digestion peut être :
-
Exclusivement
intracellulaire (protozoaires et éponges) après endocytose,
-
A
la fois intracellulaire et extracellulaire (ou luminale) comme chez les
cnidaires, les plathelminthes (turbellariés, trématodes), les mollusques
bivalves et quelques annélides (arénicole)
-
Exclusivement
luminale comme chez les nématodes, les mollusques gastéropodes et céphalopodes,
les annélides, les arthropodes, les échinodermes et les vertébrés.
Il
y a le développement de glandes digestives chez les crustacés, les arachnides
et les mollusques.
La glande
digestive est formée de tubules comprenant des cellules sécrétrices d’enzymes,
des cellules absorbantes :
-
Absorption
de petites molécules,
-
Phagocytose
de particules alimentaires.
§ Les mollusques
lamellibranches :
La glande
digestive s’ouvre sur l’estomac. Les particules triées sont digérées dans cet
organe.
§ Les crustacés :
Il y a un
déversement des enzymes dans la chambre pylorique et dans les diverticules où
s’effectue la digestion. L’absorption des nutriments s’effectue dans la glande
digestive (contraction musculaire lisse, absence de cils)
§ Les arachnides :
La digestion est
extracorporelle : les enzymes sont pompées par la région du pharynx et
proviennent de la glande digestive.
§ Les insectes :
Le tube digestif
moyen est constitué de l’intestin moyen (dilaté) et de cœca en position
antérieure. L’intestin moyen comprend des cellules sécrétrices d’enzymes et
d’absorbantes (fonctions exercées par le même type cellulaire)
Les cellules
sécrètent la membrane péritrophique (chitine et glycoprotéines) protégeant
les cellules épithéliales contre les actions mécaniques des aliments. Cette
membrane est perméable aux enzymes et aux produits de digestion.
§ Les vertébrés :
La digestion
post–pylorique des aliments est assurée par des enzymes sécrétées par le
pancréas qui déverse son suc au niveau du duodénum (rôle digestif prépondérant)
Ces enzymes ont un pH optimal voisin de 7 – 8.
Le pancréas
sécrète donc également des bicarbonates qui neutralise le chyme gastrique.
Cette sécrétion est stimulée par la sécrétine libérée par les cellules
endocrines du duodénum sous l’effet du chyme gastrique acide.
Le suc
pancréatique est déversée sous l’effet d’une hormone : la cholécystokinine
libérée par les glandes endocrines du duodénum. Cette hormone participe
également à la stimulation du déversement de la bile par la vésicule biliaire
(contraction de la vésicule biliaire)
Le
foie est un organe dont le rôle est primordial dans la physiologie de
l’organisme :
-
Fonction
glycogénique,
-
Synthèse
et dégradation des lipides,
-
Excrétion
azotée, détoxication.
Dans le cadre
des fonctions de nutrition, il produit des sels biliaires (tels que le
glycocholate et la taurocholine) Ce sont des détergents permettant l’émulsion
des lipides, favorisant ainsi l’action des lipases.
L’absorption des
produits terminaux de la dégradation (acides aminés, oses, micelles de
monoglycérides, acides gras, stérols, etc.) est assurée par l’intestin grêle.
Ce dernier présente des adaptations morphologiques permettant l’augmentation de
la surface absorbante (villosités intestinales)
L’absorption des
lipides se fait par pinocytose ou par diffusion. Il y a ensuite un remaniement
dans le REL et une association des triglycérides avec les protéines, entraînant
une exocytose vers les vaisseaux vers les vaisseaux chylifères.
L’absorption
des acides aminés et des oligopeptides se fait via un co–transport avec le Na+
(symport) :
-
Les
acides aminés essentiels ne sont pas métabolisés,
-
Les
autres acides aminés peuvent être métabolisés en d’autres acides aminés,
-
Les
oligopeptides sont hydrolysés pour obtenir des acides aminés.
Il
existe 4 catégories de transporteurs selon ce qu’ils transportent :
-
Les
diacides,
-
Les
dibasiques,
-
La
glycine, la proline et l’hydroxyproline
(Par exemple : le collagène est une
succession d’hydroxyprolines)
-
Les
autres.
Les oses sont
absorbés contre leur gradient de concentration par un co–transport de type
symport associé au Na+. Le gradient de
Na+ est maintenu par un transport
actif lié à la pompe Na+–K+ ATPasique au
niveau basale.
NB :
Les
entérocytes possèdent au niveau des microvillosités des enzymes membranaires
(disaccharidases, oligopeptidases, phosphatases alcalines)
Il y a l’absorption d’autres nutriments
comme les vitamines.
Ø Le
tube digestif postérieur :
§ Remodelage des fèces :
§ Absorption et excrétion de
l’eau et des sels :
-
Ches
les mammifères qui hébergent des bactéries, il y a E. coli qui dégrade par
fermentation les produits pas encore digérés et synthétise du mucus :
-
Gaz :
H2, CO2, déchets
métaboliques,
-
Acides
gras volatiles qui sont absorbés,
-
Apport
de vitamines par les bactéries.
-
Le
proctodéum dilaté des termites (= « panse restale ») héberge des
symbiontes (Trychonympha) qui
dégradent la cellulose et la lignine.
-
Le
cœcum intestinal hypertrophié des oiseaux et de certains mammifères herbivores
(rongeurs, chevaux, koalas, etc.) dégrade la cellulose.
v Les enzymes
digestives :
Les
enzymes digestives du tube digestif peuvent être mises en évidence :
-
Par
dosage enzymatique sur des extraits de tissus,
-
Par
détection histo–enzymatique sur des coupes de tissus par congélation en microscopie
optique ou en microscopie électronique à transmission.
-
Lorsque
l’on dispose d’anticorps spécifiques dirigés contre une enzyme, sa présence
peut être détectée sur des coupes histologiques (immunocytologie)
Les
enzymes sont classées par n° EC en fonction du type de la réaction
catalysée :
-
EC.1
= Les oxydoréductases,
-
EC.2
= Les transférases (transfert de
radicaux fonctionnels),
-
EC.3
= Les hydroxylases = enzymes digestives,
-
EC.4
= Les lyases (additions sur des
doubles liaisons),
-
EC.5
= Les isomérases,
-
EC.6
= Les ligases.
Ø Digestion
(hydrolyse) des protéines :
Il
faut casser les liaisons peptidiques. L’hydrolyse complète d’une protéine
requiert l’action successive de protéases présentant des spécificités
variées :
-
Hydrolyse
de la protéine en fragments peptidiques (endoprotéases),
-
Dégradation
des peptides à partir des extrémités (exopeptidases),
-
Hydrolyse
des petits fragments par des dipeptidases.
§ Endoprotéases :
·
Sérine–protéases
:
Elle nécessite
la présence d’une sérine dans le site catalytique. Le pH optimal est 7 – 8.
Exemples :
-
La
trypsine coupe après une lysine ou une arginine.
-
La
chymotrypsine coupe après une phénylalanine, un tryptophane ou une tyrosine.
L’enzyme
est sécrétée par le pancréas des vertébrés ou par la glande digestive des
arthropodes et des mollusques.
Ces enzymes sont synthétisées chez les
vertébrés sous forme de zymogènes inactifs qui sont activés lors de leur
libération dans la lumière du tractus digestif.
![]() Trypsine :
Trypsine :
Trypsinogène "
Trypsine
Entéropeptidase
Chymotrypsine :
Chymotrypsinogène "
Chymotrypsine + 2 dipeptides
Tryspine
Elastase :
Pro–élastase "
Elastase
Tryspine
L’élastase
hydrolyse l’élastine. Elle est sécrétée dans le pancréas des vertébrés
carnivoires.
·
Cystéine–protéases
:
Le pH optimal
est entre 2 et 7. C’est une enzyme dont une cystéine du site catalytique réagit
avec le substrat dans la réaction enzymatique.
Exemple :
Les cathepsines lysosomales : B, H
et L dont le pH optimal environne 5. Elles sont présentes chez les vertébrés et
les invertébrés.
·
Aspartate–protéases
:
Leur pH optimal
est inférieur à 6.
Exemples :
-
La
pepsine coupe après les acides aminés aromatiques (phénylalanine, tryptophane,
tyrosine) Elle est synthétisée par l’estomac des vertébrés sous forme de
pepsinogène.
Pepsinogène "
Pepsine
H+,
Pepsine
-
Les
cathepsines D et E sont présentes dans les lysosomes. La cathepsine D est une
enzyme digestive chez les hémiptères hématophages.
·
Métalloprotéases
:
Elles requièrent
un métal et sont inhibées par l’EDTA.
Exemple :
Les collagénases dégradent le collagène
chez les vertébrés carnivores. Les insectes dégradent le collagène avec
certaines sérine–protéases.
§ Exoprotéases et
exopeptidases :
Les
aminopeptidases sont cytoplasmiques et membranaires (dans les microvillosités)
Ce sont des métallo–enzymes dont le pH optimal est 8.
Les
carboxypeptidases sont des métallo–enzymes extracellulaires.
Procarboxylase "
Carboxylase
Trypsine
Les
aminopeptidases et les carboxypeptidases sont présentes chez les vertébrés et
les invertébrés.
NB :
Pour
de nombreuses enzymes, en particulier les protéases, il existe des inhibiteurs
naturels (généralement des peptides pour les protéases) Cette inhibition
prévient l’activation prématurée des protéases dans les vésicules de sécrétion.
Les inhibitions sont spécifiques de
chaque classe de protéases.
Ø Les
estérases :
Elles
hydrolysent les esters.
§ Les phosphatases
(phosphomono–estérases) :
O
││
R — O — P — OH + H2O " R —
OH +
H3PO4
│
OH
Il
existe :
-
Des
phosphatases acides (au pH optimal proche de 5) dans les lysosomes.
-
Des
phosphatases alcalines (au pH optimal proche de 9) qui sont membranaires. On
les trouve dans l’intestin des vertébrés.
Ce
sont des enzymes ubiquitaires dans le règne animal : elles agissent sur
une grande variété de substrats.
Il existe également des
phosphomono–estérases à substrat spécifique (telle que la glucose_6èase) qui sont généralement
intracellulaires.
§ Les phosphodiestérases :
La plupart
d’entre elles sont intra–cytoplasmiques et ne sont pas impliqués dans la
digestion.
·
Les
nucléases :
Ce sont les
DNases, les RNases et les nucléases non spécifiques (au pH optimal proche de 8)
Elles sont sécrétées dans le pancréas des vertébrés et dans la glande
d’invertébrés.
·
Les
exonucléases :
Il existe des
exodésoxyribonucléases (pour l’ADN) et les exoribonucléases (pour l’ARN)
ADN " ARN " Ribonucéoside " base azotée + ribose
Exoribonucléase (Ribo)nucléotidase
(Ribo)nucléosidase
ADN " dNMP " Ribonucéoside " base azotée + ribose
Exodésoxyribonucléase Désoxynucléotidase
Désoxynucléosidase
Exemple :
Les
phosphodiéstérases du venin de serpent sont constituées de
·
Les
endonucléases :
Elles sont
généralement peu spécifiques.
§ Les carboxy–estérases :
·
Les
lipases :
La
lipase gastrique est dépendante du Ca2+ et indépendante
des sels biliaires chez les vertébrés. Elle est aussi présente chez les
invertébrés.
La lipase pancréatique est dépendante
des sels biliaires (chez les vertébrés)
TAG (+ H2O) " DAG + 1 AG
TAG (+ H2O) " MAG + 2 AG
TAG (+ H2O) " Glycérol
+ 3 AG
·
Les
phospholipases :
(Poly)
Ø Hydrolyse
des glucides :
Les
glucides à hydrolyser sont :
-
Les
polymères de réserve :
-
Glycogène
(réserves cytoplasmiques)
-
Amidon
(dans les chloroplastes et les amidoblastes)
-
Insuline,
-
Laminarine
(algues)
-
Les
polymères de structure :
-
Cellulose,
-
Lignine,
-
Chitine.
§ Les polysaccharides :
·
L’amylase
:
Elle
dégrade (hydrolyse) les liaisons a 1–4 des résidus
glucose du glycogène et de l’amidon (a 1–4 et a 1–6)
Pour une dégradation complète, une
enzyme débranchante a 1–6_glucosidase est nécessaire.
L’a–amylase est
présente chez les vertébrés, dans la glande salivaire et le pancréas. Chez les
invertébrés, elle est présente dans l’estomac des mollusques et dans la glande
salivaire.
·
Les
laminarinases :
Elles sont
présentes chez les invertébrés marins phytophages ou microphages (mollusques
bivalves et gastéropodes) et chez les poissons téléostéens phytophages et
microphages.
·
La b_fructosidase :
Elle est
présente chez les insectes consommant de l’insuline (= polymère de fructose_b (2–1)) Elle
hydrolyse de polyosides de structure. Elle dégrade de la paroi squelettique des
végétaux supérieurs (= cellulose + hémicellulose + pectine + extensine (=
glycoprotéine))
·
La
cellulase :
Elle dégrade la
cellulose qui est un polymère linéaire de glucose b (1–4)
Cellulose " Cellodextrine " Cellobiose " 2 glucoses
Endoglucanase_b (1–4) Exoglucanase_b (1–4) Cellobiase_b_glucosidase
Les cellulases
sont présentes chez les mollusques herbivores, les annélides et les crustacés
xylophages. Chez les vertébrés et les insectes, la dégradation de la cellulose
est réalisée par des symbiontes.
·
Les hémicellulases
:
Elles dégradent
l’hémicellulose qui est un polymère b (1 –4) de
D_xylose et une ramification d’arabinose de glucose et de galactose.
·
Les
pectinases :
Elles dégradent
la pectine qui est un polymère de méthylgalacturonate. Elles sont présentes
chez les insectes xylophages.
·
Les
alginases :
Elles dégradent
l’acide alginique qui est polymère d’acide mannuronique. C’est un
polysaccharide de la paroi des algues. Elles sont présentes chez les mollusques
et les échinodermes.
·
L’agarase
:
Elle dégrade
l’agar–agar qui est polysaccharide sulfaté. Elle est présente chez les
mollusques.
·
La
lysosyme :
Elle dégrade la
peptidoglycane qui est le composé principal des parois bactériennes (muréine)
Elle est présente les mollusques bivalves et les vertébrés (dans la glande
salivaire)
·
Les
chitinases et les chitobiases :
Elles dégradent
la chitine qui est un polymère de N–acétylglucosamine b (1–4) Elles
sont présentes chez les protistes, les thallophytes (champignons) et les
protostomiens.
Chitine "
Chitibiose " N–acétylglucosamine
Chitinase Chitobiase
Elles
sont présentes chez les animaux se nourrissant d’arthropodes :
-
Les
vertébrés insectivores :
-
Les
oiseaux,
-
Les
reptiles (lézards),
-
Les
amphibiens (dans l’estomac et le pancréas),
-
Les
poissons (dans l’estomac, le pancréas, l’intestin)
-
Les
cnidaires (mangeurs de crustacés)
Elles
sont aussi présentes chez les espèces herbivores :
-
Les
oligochètes consommateurs de filaments mycéliens,
-
Les
gastéropodes pulmonés.
§ Les di– et trisaccharides
membranaires :
Dans
l’intestin, les métabolites ainsi absorbés sont distribués aux cellules grâce
au système circulatoire. Le système circulatoire sert également :
-
A
distribuer la chaleur dégagée par les réactions métaboliques,
-
A
distribuer les intermédiaires métaboliques d’organes à organes,
-
A
entraîner les déchets vers les organes excréteurs,
-
A
distribuer des gaz (sauf chez les insectes)
Distribution des métabolites :
-
Chez les
acœlomates :
Chez les
cnidaires, les nutriments passent facilement du gastroderme à traverser la
mésoglée vers l’épiderme.
-
Chez les
cœlomates :
Il y a un
système de transport avec un mouvement de liquide cœlomique (spimunculiens et
annélides) ou speudo–cœlomique (nématodes) Le mouvement du liquide se fait par
les mouvements du corps et/ou par la ciliature des tissus.
Le système
circulatoire est fermé ou ouvert.
v Introduction :
Pour la plupart
des êtres vivants, il y a un fonctionnement optimal de l’organisme avec un
milieu interne de composition bien définie et constante. C’est l’homéostasie du
milieu interne.
Pour
la pression osmotique, l’organisme est soit :
-
Osmoconforme
(surtout chez les invertébrés),
-
Osmorégulateur
(surtout chez les vertébrés)
Les problèmes
posés aux organismes par la régulation de leur concentration osmotique totale
et par la régulation de leur teneur en électrolytes sont généralement
considérés ensemble. Cependant, il s’agit de 2 aspects différents.
Le
problème de la régulation osmotique s’est posé lors de la conquête des milieux
saumâtres des eaux douces et surtout du milieu terrestre.
Les concentrations osmotiques peuvent
être différentes, et ceci à cause de milieux de vie différents et des plus ou
moins fortes variabilités, y compris de la disponibilité en eau.
Le métabolisme
hydrominéral consiste en l’osmorégulation et l’excrétion azotée qui permettent
une stabilité. Les minéraux sont le Na+, le Cl–, le K+, le Ca2+, le P, le S,
etc. et sont présents en des quantités variables dans les différents milieux.
Exemple : La
truite :
Elle vit soit en
eau douce (environnement hypotonique), soit en eau de mer (environnement
hypertonique)
Ø Les
différentes stratégies :
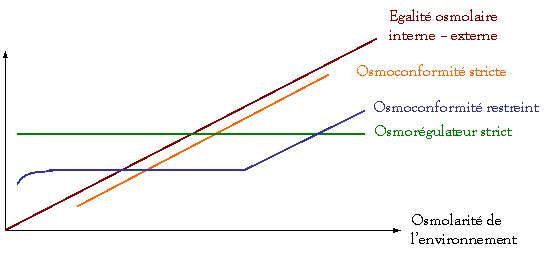
Osmorégularité
des liquides
corporels
0 10
‰ 20
‰ 30
‰
ED
< 1mmol.L–1
La mer
morte : 8 à 10 fois plus concentrée que l’eau de mer (~ 300 ‰)
Ø Les
déchets :
Les produits
terminaux du métabolisme des sucres et des graisses sont le CO2 et le H2O. Il n’y a pas
de problème particulier pour leur élimination.
Les déchets
azotés contiennent de l’azote (N) avec des groupements CHO, en plus du CO2 et du H2O.
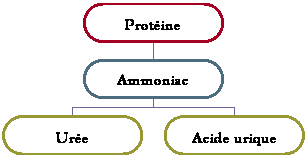
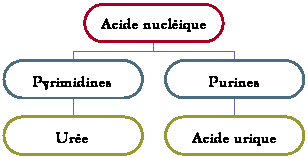
Formes des déchets :
Ammoniac :
Urée :
Acide urique :
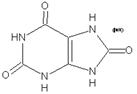
NH3/NH4+
CH4ON2
C5H4O3N4
Chez l’homme,
les déchets azotés sont constitués à 95% du catabolisme des protéines et à 5%
des acides nucléiques.
L’ammoniac est
très toxique pour l’organisme. Il peut être excrété directement par diffusion
ou après la conversion en urée ou acide urique.
Voie d’élimination = écophysiologique (=
physiologie en relation avec l’écologie) :
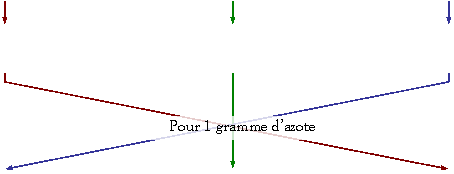 Ammoniac
Ammoniac
Urée
Acide urique
Les organismes
ammoniotéliques
Les organismes
uréotéliques
Les organismes
uricotéliques
Il faut :
500 mL d’eau 50 mL d’eau 10 mL d’eau
(Organisme
aquatique)
(Animal dans un milieu désertique)
Relation avec la notion d’économie de
l’eau :


v Osmorégulaiton en milieu
aquatique :
Ø Vertébrés
aquatiques :
Différents
mécanismes sont mis en jeu pour maintenir l’équilibre hydrominéral.
Illustration du fonctionnement
rénal :
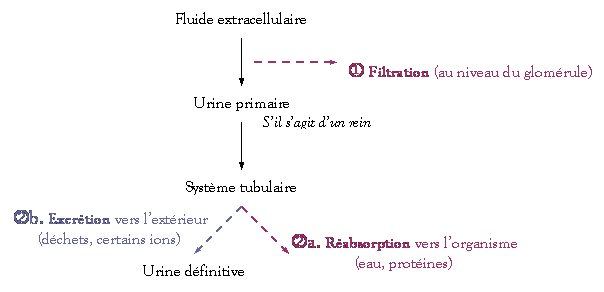
![]() Rôle du rein (mais également des branchies qui ont le même mécanisme)
Rôle du rein (mais également des branchies qui ont le même mécanisme)
Les branchies ont
un processus complémentaire : la sécrétion de mucus.
L’eau
de mer présente une osmolarité de 1000 à 1100 mOsmol.L–1 qui est
essentiellement due aux ions Na+ et Cl– :
-
Na+ : 470 mOsmol.L–1,
-
Cl– : 545 mOsmol.L–1.
" Soit environ
30g. L–1 de NaCl.
Différentes
stratégies ont été développées pour vivre dans ce milieu.
Osmoconformité globale avec des
variations importantes pour certains ions (en g.L–1) :
|
|
Na+ |
Mg+ |
Ca2+ |
K+ |
Cl– |
SO42– |
Protéines |
|
|
Eau de mer |
478,3 |
54,5 |
10,5 |
10,1 |
558,4 |
28,8 |
0 |
|
|
Méduse |
474 |
53 |
10 |
10 |
580 |
15,8 |
0,7 |
|
|
Oursin de mer (Echinus) |
476 |
54,6 |
10,5 |
10,5 |
557 |
26,5 |
0,3 |
|
|
Isopode (Ligia) |
566 |
20,2 |
34,9 |
13,3 |
629 |
4 |
0 |
|
|
langoustine (Nephrops) |
541 |
9,3 |
11,9 |
7,8 |
552 |
19,8 |
33 |
|
|
Myxine (Myxine) |
537 |
18 |
5,9 |
9,1 |
542 |
6,3 |
67 |
|
|
|
Pression
osmotique |
|
||||||
|
|
Pression
oncotique |
|||||||
§ Les cyclostomes :
Il s’agit des
myxines et les lamproies. Ces dernières peuvent aussi vivre en eau douce.
Ce sont les plus primitifs des vertébrés
vivants, ce sont des agnathes (= sans mâchoires mobiles)
Chez les
cyclostomes, l’osmolarité est similaire à celle du milieu extérieur.
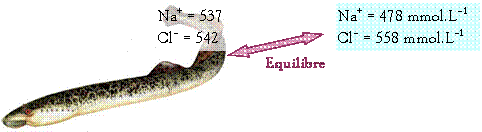
Ce sont des
espèces iso–osmotiques : il n’y a pas de problème majeur d’équilibre
hydrique. Les concentrations globales internes et externes sont similaires.
Ces espèces sont
cantonnées à des milieux où la salinité est peu variable. Ce sont des sténohalins qui ne supportent que de
très faibles variations de la concentration en sels du milieu.
Sténos = étroit ;
Halos = sels
Certaines
lamproies d’eau de mer migrent en eau douce pour la reproduction. Elles
présentent ainsi un comportement anadrome (= opposé d’un comportement
catadrome)
Elles meurent quelques temps après cette
période de reproduction.
Remarque :
Il existe aussi
des lamproies d’eau douce.
§ Les sélaciens (élasmobranches
= poissons cartilagineux) :
Les roussettes, raies et
requins :

L’équilibre
global est iso–osmotique.
Il y a un
transport actif de Na+ au niveau du
rein et de la glande rectale pour baisser la concentration interne en Na+.
La concentration sanguine en urée est
100 fois supérieure chez les sélaciens marins par rapport aux mammifères (où
l’urée est éliminée au niveau rénal)
Urée : TMAO :
=
Oxyde de triméthylamine
Chez
les sélaciens, l’urée est réabsorbée au niveau rénal.
Chez la raie
électrique (sténohalins d’eau douce), la concentration d’urée éliminée est
négligeable. La pression osmotique est de 300 mOsmol.
§ Les téléostéens :
Chez les
poissons téléostéens, l’osmolarité est € à ‚ de celle de
l’eau de mer. Les marins et les dulcicoles (= d’eau de mer) sont dans la même
fourchette.
·
En
milieu marin :

Milieu
hypertonique

Stratégie osmorégulatice :
-
Une
urine réduite et isotonique : la concentration en sels est sensiblement la
même que le sang,
-
L’excrétion
des ions par un transport actif,
-
Présence
dans les branchies de ionocytes (anciennement appelées cellules à chlorures) où
l’on trouve des pompes Na+K+ATPase.
L’urine
est concentrée, mais elle n’est pas hypertonique : le rein n’a pas d’anse
de Henlé.
Le volume
urinaire faible (0,035 mL.H–1 pour 100g de
poisson)
La réabsorption de l’eau s’effectue au
niveau de l’intestin.
·
En
eau douce :

Milieu
hypotonique
Stratégie osmorégulatice :
-
Une
urine très abondante et diluée,
-
Réabsorption
des ions par transport actif,
-
Présence
dans les branchies de ionocytes où l’on trouve des pompes Na+K+ATPase.
L’urine
éliminée en 24 H peut représenter le tiers de la masse de l’animal. Sa
concentration est de 2 à 10 mmol.L–1.
Le volume
urinaire est de 0,35 mL.H–1 pour 100g de
poisson.
La réabsorption des ions s’effectue au
niveau de l’intestin.
§ Récapitulatif des
vertébrés :
Osmolarité
externe
(en mOsmol.kg–1 de H2O)
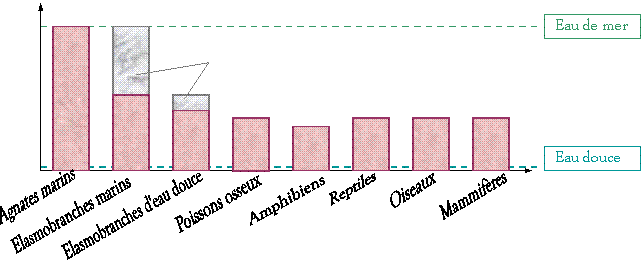
1000
Composés d’urée OTMA
500
0
Ø Invertébrés
aquatiques :
La
plupart des invertébrés présentent un milieu intérieur de concentration
osmotique similaire à celle de l’extérieur. Ce sont des osmoconformes.
La pression osmotique des invertébrés marins
varie en fonction de celle de l’eau de mer.
Exemple :
L’artémia :
C’est
un crustacé des grands lacs des USA dont la salinité peut atteindre 300 ‰.
L’artémia peut
se comporter en osmoconforme, mais aussi en osmorégulateur (via des transports
de NaCl au niveau branchial)
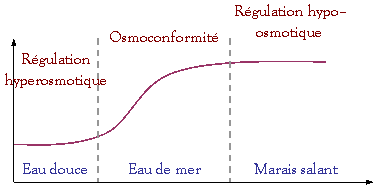 Potentiel
osmotique
Potentiel
osmotique
des fluides
corporels
Potentiel
osmotique
de
l’environnement
v L’excrétion azotée et
l’osmorégulation :
Ø Les
différentes voies (rappel) :
La régulation du
métabolisme hydrominéral consiste en une déperdition d’eau plus ou moins
importante qui est mise au profit de l’élimination de divers déchets, dont les
déchets azotés.
Chez
les invertébrés, l’excrétion azotée d’effectue
fréquemment par diffusion passive de l’ammoniaque dans l’eau.
L’urée est élaborée à partir de
l’ammoniac et du CO2 via le cycle de
l’urée (hépatique)
Ø Les
systèmes excréteurs des invertébrés :
§ Les vésicules
pulsatiles :
Ce ne sont pas
des organes mais des organites. C’est la 1ère forme spécialisée dans
la régulation osmotique (chez les protozoaires et les spongiaires d’eau douce)
Elles
permettent des transports actifs de Na+ à partir de
microvésicules qui fusionnent puis s’ouvrent à la périphérie de la cellule et
déversent leur contenu à l’extérieur. Un nouveau cycle peut alors recommencer :
-
Le
contenu intravésical est iso–osmotique par rapport au cytoplasme,
-
Il
y a transport actif via des NAK–ATPase : le contenu intravésical devient
hypo–osmotique,
-
La
microvésicule fusionne avec la vacuole et y déverse son contenu,
-
La
vacuole expulse le liquide à l’extérieur par contraction.
§ Les néphridies :
C’est la forme
la plus primitive des organes d’excrétion et d’osmorégulation. Le principe de
base est la filtration des liquides extracellulaires accompagnée de
réabsorption ou d’excrétion actives de certains ions (ou molécules)
·
Les
protonéphridies :
Protos = avant Néphros = rein
Ce sont des
structures tubulaires et le plus souvent ramifiées.
Une extrémité est « aveugle »
(c’est–à–dire fermée), l’autre s’ouvre vers l’extérieur via un pore : le
néphridiopore.
Ce
sont des structures classiques chez les acœlomates tels que les plathelminthes.
Exemple d’un planaire d’eau douce :
Un
réseau de tubules se terminent par des cellules spécialisées : les
cellules flammes. L’entrée des fluides corporels se fait au niveau de ces
cellules. Ils sont ensuite acheminés par les tubules vers le pore excréteur
grâce aux battements des cils qui créent un courant.
Il y a la production d’un fluide
hypotonique (= « urine ») avec une réabsorption assurée au niveau
tubulaire.
Ce système est
présent chez les rotifères, plathelminthes (embranchement des vers plats) et
les larves de mollusques.
·
Les
cœlomoductes :
L’apparition de
l’appareil du coelome est une étape importante dans l’évolution des
métazoaires. Le compartiment cœlomique est une unité anatomique autour de
laquelle s’organise la physiologie du l’animal (formation des appareils à
vocation collectives : digestives, circulatoires, etc.)
Chez
les formes primitives, la fonction excrétrice reste
« régionalisée » avec des cœlomoductes métamérisés (=
métanéphridies)
Chez les annélides, chaque métamère est
une paire d’organes excréteurs (ou métanéphridies) faisant communiquer la
cavité cœlomique et le milieu extérieur.
Il
y a la présence d’ouvertures internes qui recueillent les liquides biologiques.
De plus, les vaisseaux sanguins sont étroitement associés aux tubules
excréteurs.
A l’une des
extrémités, il y a le néphrostome. C’est un entonnoir cilié qui recueille le
liquide cœlomique provenant du segment corporel antérieur voisin.
A l’autre extrémité, le tubule se vide à
l’extérieur par le néphridiopore.
Il
y a réabsorption des électrolytes créant une urine hypotonique. Cette urine
diluée peut représenter 60% de la masse du ver. Les métanéphridies compensent
l’osmose continue se produisant à travers la peau de l’animal dans un sol
humide.
Exemple d’une espèce terrestre qui se
comporte ici comme une espèce aquatique.
§ La transformation des
cœlomoductes excréteurs en organes massifs : le rein des mollusques :
La
plupart des mollusques marins excrètent leurs déchets azotés essentiellement
sous forme d’ammoniac (50%) via une diffusion à travers l’épithélium branchial.
Le rein est
impliqué dans l’excrétion d’autres substances azotées : acides aminés,
OTMA. Il joue un rôle peu important dans l’osmorégulation (isotonicité par
rapport au milieu extérieur)
Dans le cas des espèces continentales,
il joue un rôle dans l’économie hydrique (excrétion d’urée et d’acide urique)
§ Les tubes de Malpighi des
insectes :
C’est
l’appareil excréteur majeur des arthropodes terrestres. Il est considéré comme
une extension (évagination) tubulaire de la partie distale du tube digestif.
 Ces tubes
baignent dans l’hémolymphe, régulent sa composition ionique, extraient les
composés azotés (leur nombre varie de 2 à plusieurs centaines)
Ces tubes
baignent dans l’hémolymphe, régulent sa composition ionique, extraient les
composés azotés (leur nombre varie de 2 à plusieurs centaines)
Intestin
moyen
Rectum
Tube
de Malpighi
Parfois, il
existe des reins dits « d’accumulation » (chez les arthropodes
terrestres) Le rejet des catabolites azotés ne se produit qu’à la mort de l’animal.
Exemple particulier : la glande verte des crustacés (crabe,
écrevisse, homard)
Elle
remplace les tubules de Malpighi. C’est un sac terminal relié à la vessie par
un canal néphridien. Les pores excréteurs sont localisés sous chaque antenne.
La glande baigne dans l’hémolymphe.
Le filtrat formé
au niveau du sac terminal passe par le canal néphridien, puis dans la vessie et
est éliminé au niveau des pores excréteurs.
Il
existe des différences de structures et de fonctions entre les glandes vertes
d’écrevisses (eau douce) et celles des langoustes (eau de mer)
Eau douce : il y a une
quantité importante d’urine hypotonique (conservation des sels) Il y a donc un
transport actif des sels vers le cœlome et la sécrétion dans le canal des
molécules à éliminer.
Eau de mer : le canal
néphridien est plus court (pas besoin de conserver les sels)
§ Le néphron des
vertébrés :
§ Exemple d’autres organes
spécialisés :
·
Les
branchies des poissons téléostéens :
Ce
sont des cellules respiratoires, à mucus, ionocytes (= cellules à chlorures)
Elles sont riches en mitochondries.
Elles prolifèrent lors du passage en eau
de mer et se différencient (augmentation de la taille, de l’activité
NaK–ATPase)
In vitro, des hormones telles que le cortisol,
·
Les
glandes à sel :
Elles sont
principalement annexées à la région cloacale de certains poissons, à l’œil ou
l’organe olfactif des reptiles marins, des reptiles désertiques et des oiseaux
marins.
Elles permettent
la sécrétion d’un liquide aussi (voire plus) concentré en sels que l’eau de
mer. Cette sécrétion intervient surtout après un stress osmotique (ingestion
d’eau de mer ou de nourriture salée)
 Na+ excrété
Na+ excrété
(mmol)
Excrétion
nasale
Excrétion
cloacale
Temps
(min)
L’élimination
nasale représente 43,7 mmoles au total contre 4,41 mmoles pour ce qui est de
l’élimination cloacale. Et cela, malgré un volume plus faible 56,3 mL (volume
d’élimination nasale) contre 75,2 mL (volume d’élimination cloacale)
Exemples de la glande rectale des
sélaciens et de la glande lacrymale des tortues marines ou des crocodiles.
§ Le rein :
Il
est impliqué dans de nombreuses fonctions :
-
La
régulation de l’osmolarité et du volume des liquides extra–cellulaires,
-
La
régulation de l’équilibre des électrolytes,
-
La
régulation de l’équilibre acido–basique,
-
L’excrétion
des déchets et des substances étrangères,
-
La
production et la sécrétion d’hormones, comme :
-
La
rénine dans le système rénine–angiostensine–aldostérone (pression artérielle),
-
Le
1,25 (OH) 2D3 (= vitamine D)
Le néphron peut
être court ou long, avec une structure en « épingle à cheveux ») Cela
concerne le néphron des vertébrés (800 000 à 120 000 néphrons par
rein)
·
Filtration
glomérulaire :
La filtration
s’effectue dans une « capsule creuse » en forme de ballon : la
capsule de Bowman. Le glomérule est constitué de nombreux capillaires reliés
entre–eux (= capillaires gloméruleux)
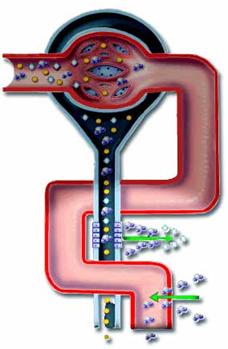
La majorité des
molécules du sang sont filtrés.
·
Réabsorption
tubulaire :
Valeurs moyennes de plusieurs composants
traités par filtration et réabsorption :
|
Substance |
Quantité
filtrée par jour |
Quantité
excrétée par jour |
Pourcentage
réabsorbé |
|
Eau (en L) |
180 |
1,8 |
99 |
|
Na+ (en g) |
630 |
3,8 |
99,5 |
|
Glucose (en g) |
180 |
0 |
100 |
|
Urée (en g) |
54 |
30 |
44 |
Le début de la
réabsorption se fait dans le tubule proximal. Dès que l’urine primitive atteint
50% de la longueur du tube proximal du néphron, l’urine devient 2,5 fois plus
concentrée que le plasma, témoin d’une réabsorption importante d’eau couplée à
la réabsorption active de Na+.
Quantité de
glucose
 (mg/min)
(mg/min)
Glucose
réabsorbé
Glucose
excrété
![]() Quantité de glucose
Quantité de glucose
filtré
(mg/min)
Quantité
physiologique
Il y a
saturation du système de transport du glucose. Le glucose administré a un sujet
par la voie intraveineuse, si bien que le glucose plasmatique et, par
conséquent, le glucose filtré augmentent.
180 L/jour Û 180 L/jour Û 7,5 L/heure Û 7 g/heure Û 110 mg/min
Il existe une
relation entre la longueur de l’anse de Henlé et la capacité de concentration
de l’urine. Par exemple, chez les poissons, il n’y a pas d’anse de Henlé et
l’urine ne peut pas être concentrée que le plasma.
|
Animal : |
Concentration de l’urine (en mOsmol.L–1) |
Rapport de concentration de l’urine sur celle du
plasma |
|
Castor (vit
dans l’eau) |
520 |
2 |
|
Homme |
1400 |
4 |
|
Rat blanc =
consommateur de graines (peu hydratée) |
2900 |
9 |
|
Chat |
3100 |
10 |
|
Rat du désert |
6300 |
17 |
v Les échanges calciques
(homéostasie) :
Calcémie :
-
Paramètre
stable chez de nombreuses espèces (2,2 à 2,6 mmoles.L–1) : les
vertébrés mammaliens, les oiseaux, les reptiles et les amphibiens.
-
Chez
les vertébrés aquatiques, il y a plusieurs voies d’entrée pour éviter
l’hypercalcémie.
-
Chez
les vertébrés aériens, il y a des apports discontinus pour éviter
l’hypocalcémie.
Chez
les poissons téléostéens et amphibiens aquatiques :
-
Il
y a une réserve de calcium.
-
Il
existe des corps ultimobranchiaux sécrétant la calcitonine (hypocalcémiante)
-
Il
existe des corpuscules de Stannius sécrétant la stanniocalcine (hypocalcémiante)
-
La
vitamine C est hypercalcémiante.
Chez
les tétrapodes aériens (passage à la vie terrestre) :
-
Il
y a
-
Les
cellules C de la thyroïde proviennent des CUB et sécrètent la calcitonine qui
est hypocalcémiante (chez les mammifères,
oiseaux, reptiles et amphibiens terrestres)
-
Il
existe 3 hormones majeures pour 3 effecteurs majeurs.
Echanges calciques (chez l’homme) :
Au
niveau de l’intestin, sur
-
350
mg sont absorbés par l’organisme,
-
150
mg reviennent dans l’intestin.
Au niveau des
fèces : 1000 – 350 + 150 = 800 mg.
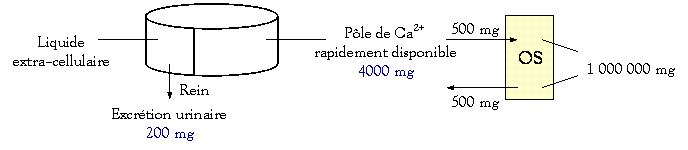
v Calcémie et sa régulation
(chez les vertébrés aériens) :
Ø Coté
hormonal :
§ La vitamine D :
Il y a 2 sources
de vitamines D : l’alimentation et le 7–déshydroxycholestérol = précurseur
qui peut se transformer en vitamine D sous la peau grâce aux rayons du soleil.
La
vitamine D passe ensuite dans le foie, où elle est hydroxylée en 25–OH vitamine
D pour arriver dans les reins :
-
En
cas d’excès de Ca2+, la vitamine D
est hydroxylée en 24,25–(OH)2 vitamine D
(vitamine D inactive, donc hypocalcémiante)
-
En
cas de déficience de Ca2+, la vitamine D
est hydroxylée en 1,25–(OH)2 vitamine D
(vitamine D active, donc hypercalcémiante)
Contrôle de la production rénale des
métabolites dihydroxylés de la vitamine D3 par la calcémie chez le rat :
24,25–(OH)2 D
1,25–(OH)2 D
Calcémie (mg.L–1)
§ La PTH (parathormone) :
Contrôle de la sécrétion de PTH par la
calcémie chez le jeune veau :
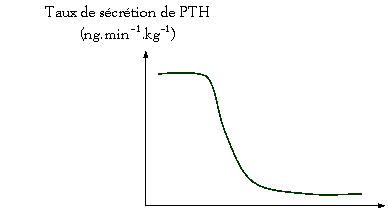
Calcémie
(mg.L–1)
§ La calcitonine :
Corrélation entre la calcémie et le taux
se sécrétion de calcitonine chez le fœtus de mouton à terme :
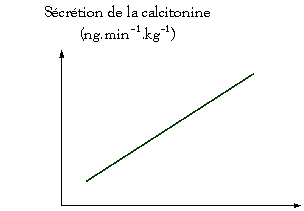
Chargement
de la calcémie
Ø Coté
effecteurs :
§ L’intestin :
Il est impliqué
dans la réabsorption du Ca2+.
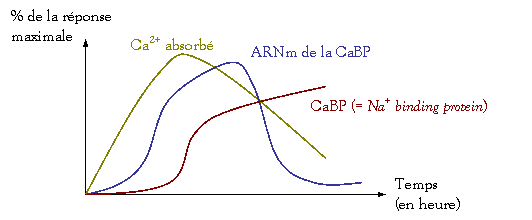
La vitamine D
agit en augmentant la quantité de CaBP.
§ L’os :
Il
est en perpétuelle régénération : il y a un équilibre entre construction
et dégradation.
Les ostéoblastes
fabriquent de nouveaux tissus en utilisant le Ca2+ sanguin. Ils
sont aussi impliqués dans la dégradation de l’os et la libération du Ca2+.
La calcitonine a
un effet inverse de
§ Le rein :
La
calcitonine a un effet négatif sur la réabsorption du Ca2+.
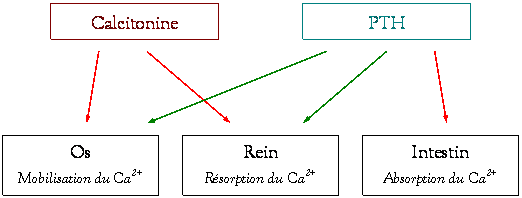
Ø Exemple
de type de réulation :


 ( [Ca2+] plasmatique
( [Ca2+] plasmatique
ß
& Sécrétion de
PTH
ß
& [1,25(OH)2
D]
ß
& Absorption
intestinale
&
Résorption de
l’os
ß
![]() &
[Ca2+] plasmatique Si
excès
&
[Ca2+] plasmatique Si
excès
![]() ( [Ca2+] urinaire
( [Ca2+] urinaire
& Sécrétion de
calcitonine
Ø Remarque
et fin :
Chez les
poissons, il existe d’autres hormones : stanniocalcine (STC) qui inhibe de
l’entrée de Ca2+ et qui est
stimulée par la prolactine.
La respiration
se réfère aux processus biochimiques qui permettent la libération d’énergie à
partir des métabolites (essentiellement des oses) Ces processus sont
essentiellement (bien que non exclusivement) aérobiques. On parle de
« respiration interne » ou « respiration tissulaire ».
La
respiration peut également se référer aux processus permettant l’obtention d’O2 (depuis
l’environnement) et le rejet de CO2 produit par la
respiration tissulaire. Ce 2nd aspect physiologique de la
respiration est souvent qualifiée de « respiration externe ».
Cette activité physiologique
(respiration externe) requiert les sites d’échange entre l’organisme et le
milieu environnant, mais également un système circulatoire (si on exclue les
animaux de petite taille) qui distribue l’O2 aux tissus. Le
système circulatoire sert à la distribution des nutriments et des déchets en
particulier chez les insectes.
v Systèmes
respiratoires :
Ø Structure
des principaux systèmes respiratoires :
Les molécules
d’oxygènes diffusent du milieu à forte PO2 vers le milieu
à faible PO2 (pression partielle en O2) La pression
totale d’un mélange gazeux est la somme des pressions partielles des gaz le
constituant.
PO2
= FO2 x P
F
= fraction d’O2 dans le mélange
P
= pression totale du mélange
A l’équilibre,
les PO2 sont égales entre les 2 milieux
d’échange. Cependant, 2 milieux en équilibre n’ont pas nécessairement la même
concentration moléculaire en gaz. Ceci dépend du coefficient de solubilité de
ce gaz dans ce milieu.
Le
taux de diffusion d’un gaz dans les tissus dépend du gradient de pression de ce
gaz et aussi des propriétés du tissu (exprimées par le coefficient de
diffusion)
On estime que la
distance maximale de diffusion ne peut donc concerner que :
-
Les
animaux de petite taille (protozoaires),
-
Les
plathelminthes qui présentent une forme aplatie,
-
Les
nématodes (tubes allongés), les éponges, les coraux (surfaces complexes),
-
Les
organismes qui ont un métabolisme très bas comme les méduses qui, malgré leur
grande taille, ne sont constituées que d’1% de matière organique (leurs
cellules actives sont donc localisées en surface)
Pour les animaux
de grande taille : si on multiplie la surface par 4, il faut multiplier le
volume par 8. Il faut donc la mise en place de systèmes spécialisés
d’échangeurs de gaz.
§ Les branchies :
Ce sont des évaginations
corporelles vers l’extérieur, qui baignent à l’extérieur de l’animal mais qui
peuvent être enfermées dans une cavité comme chez les poissons, ou chez les
mollusques (dans la cavité palléale)
·
Branchies
simples :
Ce sont des
évaginations simples de la peau rencontrées chez certains échinodermes (oursins
et étoile de mer) Elles peuvent être segmentées sous forme de touffe comme chez
de nombreux annélides aquatiques.
·
Branchies
filamenteuses :
Elles
ont une structure en forme de plume chez les arthropodes aquatiques, recouverts
d’une fine couche de cuticule externe avec un système circulatoire branchial
plus élaboré.
Cette structure est également observée
chez certaines salamandres, têtards et au niveau des nageoires pelviennes de
certains dipneustes.
·
Branchies
lamellaires :
Ce
sont des extensions aplaties d’un pilier en arc branchial. On les retrouve
chez :
-
Les
pléopodes (appendices modifiés) des isopodes,
-
Les
décapodes : expansion également à la base des appendices. Les espèces
littorales peuvent s’en servir pour la respiration aérienne.
-
Les
élasmobranches et les poissons téléostéens (jusqu’à 14 paires de branchies
protégées par un opercule)
NB :
En général, les
branchies sont peu adaptées pour la respiration aérienne : les filaments
s’affaissent sur eux–mêmes et s’accolent. Cela concerne les espèces ayant
conquis le milieu aérien récemment.
Exemples :
-
Le
crabe des cocotiers présente des branchies rigides pour lesquelles il a besoin
d’un taux d’humidité importante.
-
Les
cloportes (isopodes) présentent des branchies à l’intérieur de cavités (= sorte
de poumons fonctionnels)
-
Chez
les arachnides (scorpions, araignées), l’organe respiratoire ressemble à des
branchies enfermées dans une chambre. Les lamelles sont maintenues
indépendantes par des piliers articulaires (appelés également poumons
lamellaires)
NB :
La différence
entre branchie et poumons est que les branchies sont des extensions externes
alors que les poumons sont des replis internes.
§ Les poumons :
Si la structure
corporelle s’invagine, la cavité créée est appelée poumon.
Si cette structure peut se rencontrer
chez quelques animaux marins comme chez les concombres de mer
(échinodermes/holothuries), les poumons sont généralement rencontrées chez les
espèces terrestres pour lesquelles des structures superficielles comme les
branchies représenteraient un désavantage compte–tenu des pertes en eau.
Les
poumons peuvent être de simples sacs comme chez certains crabes terrestre chez
lesquels les branchies lamellaires sont absentes mais la chambre branchiale est
richement vascularisée.
Ces simples poumons se rencontrent chez
les mollusques gastéropodes pulmonés (escargots, limnées) La chambre résulte du
repliement de manteau qui est richement vascularisé. La ventilation est assurée
par les mouvements du pied.
Chez les
poissons, la respiration est faite par une vessie natatoire (poissons
primitifs), ou une poche suprabranchiale ou encore une partie du tube digestif.
Les
véritables poumons sont rencontrés chez les tétrapodes : amphibiens,
reptiles, oiseaux et mammifères. On y observe une complexité croissante.
Chez les oiseaux
et les mammifères, l’endothermie requiert une consommation d’O2 accrue. Pour
assurer cet apport, il y a une augmentation de la surface pulmonaire par
la mise en place d’un réseau dense de bronchioles et d’alvéoles (1 cm3 de poumon de
mammifère correspond à 600 cm2 de surface) Le
volume des poumons est proportionnel à la masse de l’animal.
Chez les
oiseaux, des poumons compacts de petite taille communiquent avec de volumineux
sacs aériens. Les ramifications très fines des bronches ne se terminent pas en
cul de sac comme chez les mammifères (absence d’alvéoles), mais au contraire
permettent un passage continu de l’air via des parabronches qui communiquent
avec un réseau anastomosé de capillaires aériens.
§ Le système trachéen des
insectes et de certains arthropodes terrestres :
Ce système de
tubules qui conduit l’air aux tissus existe chez les insectes et quelques
arthropodes terrestres (myriapodes, nombreux arachnides, millepattes)
NB :
Dans l’air, l’O2 diffuse environ
300 000 fois plus vite que dans l’eau. Certains crustacés isopodes
terrestres possèdent des branchies et des trachées dans leurs pléiopodes
abdominaux (pseudotrachées)
·
Insectes
ptérygotes :
Ils présentent
une série de stigmates qui donnent sur une trachée IAIRE qui se ramifie
en trachées IIAIRES qui conduisent
à des trachées IIIAIRES qui se
ramifient dans les tissus pour atteindre un diamètre de 0,2µm (trachéoles)
Il y a la
présence de sacs aériens chez les insectes volants. Ces sacs peuvent être
comprimés et ainsi entraîner une modification du vol par le mouvement du corps.
L’extrémité
des trachées contient du liquide en plus ou moins grande quantité, permettant
une régulation de la diffusion de l’O2 dans les
cellules.
Si l’activation métabolique est intense,
les réserves sont dégradées en petites molécules, entraînant une augmentation
de la pression osmotique des cellules et donc de l’absorption d’eau, ce qui
provoque la diffusion d’O2.
·
Modification
chez les insectes aquatiques :
Soit :
Ils présentent
un système clos, sans stigmates. La diffusion des gaz se fait à travers la
cuticule, puis il y a une distribution de l’O2 dans le système
trachéen.
Soit :
Ils présentent
des expansions abdominales, des sortes de branchies (= trachéobranchies) qui
permettent des échanges entre l’eau et le système trachéen. C’est le cas chez
les larves d’éphémères.
Soit :
Les
trachéobranchies sont localisées dans la lumière du rectum et permettent
également la nage par réaction.
NB :
Certains
insectes aquatiques emprisonnent une bulle d’air. Le dytique emprisonne de
l’air sous ses ailes. Les échanges se font par diffusion.
Il y a un
équilibre entre les pressions totales de l’eau et de l’atmosphère. A 1m de
profondeur, la pression totale augmente, les pressions partielles aussi. Cela
entraîne une perte d’O2 et de N2.
Lors d’une activité, l’O2 est consommé si
bien que la pression partielle en O2 de la bulle devient
inférieure à celle de l’eau. La pression partielle en N2 de la bulle est
supérieure à celle de l’eau, donc l’azote diffuse vers l’eau et sort de la
bulle.
D’autres
insectes aquatiques (comme les scarabées) possèdent un plastron constitué de
poils hydrofuges, emprisonnant de l’air :
-
La
bulle est incompressible, il n’y adonc pas de contraction de la bulle sous la
pression.
-
L’O2 diffuse de
manière continue dans l’air emprisonné.
-
La
pression partielle en N2 reste en
équilibre avec l’eau.
§ La peau :
De nombreux
animaux ne possèdent pas de structures spéciales d’échangeur de gaz et absorbent
la majorité de leur O2 par la peau.
Ceci
est possible s’ils possèdent un système efficient de perfusion permettant au
sang désoxygéné d’irriguer la surface respiratoire. Cela concerne :
-
Les
petits animaux aquatiques, les larves et les vers,
-
Les
annélides semi–terrestre (vers de terre) et mollusques aquatiques tropicaux
(limaces tropicales),
-
Quelques
vertébrés aquatiques comme les 1ers stades larvaires de poissons,
les anguilles adultes, les salamandres, les serpents marins et également chez
la plupart des amphibiens terrestres anoures (grenouilles et crapauds)
Chez ces
vertébrés, ceci semble être un développement secondaire dans la mesure où les
espèces ancêtres immédiates respirent avec des branchies ou des poumons.
Ø Mécanismes
de ventilation permettant une augmentation des échanges gazeux :
§ Mécanismes :
Les barrières
d’échanges gazeux des systèmes respiratoires sont très fines : souvent une
couche de cellules épithéliales + du mucus ou de la chitine (donc une épaisseur
souvent inférieure au µm)
Le
renouvellement du milieu en contact avec la surface respiratoire est fait par
la création d’un flux de fluide qui accroît le taux de diffusion (Q) :
Q = F
(C2–C1)
La création de se flux requiert
cependant de l’énergie.
-
Le
mouvement de flagelles de choanocytes des éponges assure la ventilation mais
également la filtration.
-
Le
mouvement ciliaire assure un courant d’eau au niveau des branchies des
mollusques bivalves (ventilation et filtration)
NB :
Chez les
mollusques bivalves, l’importance des branchies dans la respiration n’est
peut–être pas primordiale, la surface du manteau pourrait suffire à assurer les
échanges gazeux nécessaires (animaux peu actifs)
Le rôle des muscles :
La
ventilation peut être associée à la locomotion comme chez :
-
Les
céphalopodes (nage à réaction),
-
De
nombreux poissons (thon, requins, maqueraux) qui ont besoin d’une ventilation
par flux continu,
Chez les téléostéens :
Le
mouvement se fait par pompage.
-
L’augmentation
du volume de la cavité buccale se fait par un abaissement du plancher buccal.
Il y a la fermeture de la bouche, créant une augmentation de la pression. Le
flux d’eau est dirigé vers la cavité operculaire.
-
Le
volume de la cavité operculaire peut être accru par le mouvement des opercules,
faisant sortir l’eau.
La bouche et les
opercules fonctionnent comme des valves permettant ainsi un flux
unidirectionnel (mécanisme analogue de la ventilation pulmonaire chez les
batraciens qui avalent l’air. La fréquence de pompage est fonction de la
concentration en O2.
Chez les crustacés décapodes (crabes,
homards) :
La
présence d’une « palme » (ou scaphognathite) attachée aux maxillaires
permet le pompage de l’eau dans la chambre branchiale.
Ce dernier est également capable
d’inverser brièvement son sens de battement, ce qui provoque ainsi le nettoyage
de la cavité branchiale. La fréquence des battements est fonction de la concentration.
Chez les crustacés isopodes (cloportes,
idotée) :
La ventilation
est assurée par le battement des pléopodes (= appareils abdominaux)
Chez les branchiopodes (daphnies,
arthemia) :
La ventilation
est permise par le battement des appendices thoraciques.
Chez les « gros » insectes :
La ventilation
peut être assurée par la contraction des muscles du vol qui compriment les sacs
aériens.
·
Ventilation
par aspiration par création d’une pression négative dans les poumons :
-
Par
mouvement du manteau et du pied chez les mollusques pulmonés (escargot)
-
Par
les côtes et les muscles intercostaux qui, chez les reptiles, provoquent une
expansion de la cage thoracique. Il n’y a pas de diaphragme, c’est le foie qui
joue ce rôle chez les crocodiliens.
-
Par
les côtes et le diaphragme musculaire isolant ainsi l’espace thoracique et
augmentant ainsi la pression négative, chez les oiseaux et les mammifères.
·
Chez
les oiseaux :
-
L’abaissement
du sternum conduit à une expansion des sacs aériens. L’air est conduit à
travers la mésobronche vers les sacs postérieurs.
-
L’élévation
du sternum provoque la compression de l’air des sacs postérieurs et son
transfert vers les parabronches.
-
Au
cours de la 2nde inspiration, l’air des parabronches est conduit
dans les sacs antérieurs, alors que les sacs postérieurs se remplissent d’air
provenant de l’extérieur.
-
Au
cours de la 2nde expiration, l’air des sacs antérieurs est expulsé.
§ Flux d’échange de gaz :
Les
systèmes de ventilation peuvent se caractériser par l’orientation relative du
flux du milieu externe par rapport aux flux du liquide perfusé (sang) :
-
Flux
parallèle : « co–courant » = courants convergents,
-
Flux
opposés : contre–courant »,
-
Flux
croisés : courant croisés ».
v Système circulatoire :
Le système
circulatoire permet le mouvement de fluide et entraîne la réduction des
distances de diffusion de gaz, métabolites, déchets, catabolites, NH4+, urée,
créatine, hormones, etc.
Il peut aussi avoir un rôle
hydraulique (hydrosquelette) dans la locomotion ou l’érection.
Ø Plans
d’organisation :
-
Pompage
du milieu externe : éponges, cnidaires.
-
Circulation
de liquide interne (système circulatoire)
Elle consiste en le mouvement du liquide
extracellulaire. Le liquide circule à travers des sinus dans les tissus et dans
les cavités aux lacunes plus larges entre les tissus.
Le liquide
s’appelle hémolymphe. Le système circulatoire est de type ouvert ou lacunaire.
-
Les
petits animaux acoelomates et pseudocœlomates (plathelminthes, némathelminthes)
Le liquide du blastocèle baigne les tissus. Le mouvement du corps assure la
circulation.
-
Chez
les cœlomates (arthropodes/mollusques), l’hémolymphe est partiellement
véhiculée par des vaisseaux sanguins qui peuvent percoler certains organes
(branchies, organes excréteurs) Elle passe dans l’hémocèle qui baigne les
autres tissus et retourne dans les vaisseaux grâce à un ou plusieurs cœurs (=
vaisseaux plus épais et plus musculeux)
NB :
Le volume du
liquide est important (jusqu’à 30% de la masse corporelle) présentant une
faible pression, avec des temps longs de circulation.
Ø Le
fluide circulant peut être séparé du liquide extracellulaire :
Le fluide
circulant est le sang, le système circulatoire est clos. Le sang traverse des
vaisseaux délimités par des cellules endothéliales. Le sang entre en contact
avec les tissus quand il traverse la paroi des capillaires.
Ce système
circulatoire nécessité des pompes puissantes (cœurs) Le liquide constitue un
faible volume (environ 8% de la masse corporelle chez les mammifères) La
pression artérielle est importante. Sa composition est plus stable.
Il est observé
chez les mollusques céphalopodes (présence de 3 cœurs) Chez les annélides
polychètes et oligochètes, le système circulatoire est clos. Cependant,
certains annélides possèdent des régions délimitées par les tissus à travers
lesquelles le sang circule. Certaines sangsues ont un système circulatoire
ouvert.
Les
vertébrés :
Le cœur
correspond à une section d’un vaisseau sanguin principal épaissi, plus
musculeux, avec une synchronisation nerveuse et la présence de valves pour
assurer un flux unidirectionnel. Les cœurs sont mutlichambres.
Les invertébrés :
Annélides :
Certaines
sections du vaisseau dorsal sont épaissies et montrent une contraction
rythmique assurant un flux vers la région antérieure.
Chez certains oligochètes (comme le ver
de terre), les « cœurs dorsaux » sont supplémentés, voire supplantés,
par une série de cœurs pairs sur les vaisseaux latéraux qui lient les
longitudinaux ventraux et dorsaux.
Mollusques :
Ils
présentent une grande diversité. Leur cœur a 2 chambres : une oreillette
(= atrium) et un ventricule.
Le cœur est renfermé dans la cavité
péricardique assez rigide, ainsi la contraction du ventricule réduit la
pression de la cavité péricardique, ce qui accélère le retour du sang veineux
dans l’oreillette.
Chez les
céphalopodes, il y a un cœur possédant la même organisation, mais il est plus
large, plus puissant, et supplémenté par 2 cœurs branchiaux qui génèrent une
surpression de sang veineux entrant dans la branchie.
Arthropodes :
Compte–tenu
de l’absence de veines pour le retour du sang dans le cœur, le sang est
collecté par le cœur au niveau des ostioles munis de valves.
Au cours de l’expansion cardiaque
(diastole), le sang entre dans le cœur. La contraction du cœur (les valves des
ostioles étant fermées) expulse le sang dans les artères.
La
pression négative créée dans la cavité péricardique (rigide) aspire le sang
veineux. La cavité péricardique est donc un sinus sanguin et non une cavité
cœlomique.
Un système similaire est observé chez
les insectes : il est constitué d’une série de chambres cardiaques. Il
correspond au vaisseau dorsal de l’arrière de l’abdomen jusqu’au mésothorax.
Les muscles
aliformes se fixent latéralement à la paroi du cœur et au cœur par des
filaments tenseurs. Par leur contraction ordonnée, ces muscles refoulent le
sang de l’arrière vers l’avant (vers l’aorte)
Les poissons
téléostéens :
Le sinus veineux
et l’oreillette sont faiblement contractiles et peu musculeux. Le ventricule
très musculeux et fortement contractile. Le bulbe artériel est élastique.
Les amphibiens :
Leur cœur
présente une séparation anatomique incomplète des 2 circulations, mais il y a
une séparation fonctionnelle. Il y a un ventricule unique, mais 2 oreillettes
parfaitement individualisées : l’oreillette gauche reçoit le sang hématosé
et l’oreillette droite reçoit le sang veineux.
La valvule
spirale limite le mélange des 2 sangs et permet le mécanisme de stratification
lors du remplissage du ventricule par les oreillettes.
En plongée,
l’augmentation de la résistance pulmonaire entraîne dérivation du flux sanguin
vers la circulation système et cutanée et favorise les échanges respiratoires
cutanés.
Les crocodiliens :
Leur cœur
présente 2 ventricules et 2 oreillettes mais il y a une communication entre les
2 circulations au niveau des 2 crosses aortiques : la séparation n’est pas
complète.
Comparaisons des systèmes circulatoires
entre les vertébrés et invertébrés :
-
Poissons :
Circulation de
gauche à droite,
Forte pression
entre les branchies et les organes,
Très faible
pression entre les organes et le cœur.
-
Reptiles :
Faible pression
entre les organes et le cœur, et entre les poumons et le cœur,
Circulation de
gauche à droite.
-
Mammifères :
Faible
pression entre les organes et le cœur, et entre les poumons et le cœur.
NB :
En association avec
l’évolution des systèmes respiratoires, la pression artérielle a augmenté,
entraînant d’avantage de liquide et de protéines plasmatique qui sont perdues
au niveau des capillaires dans la fente interstitielle.
Ainsi, il y a
l’apparition d’un système lymphatique qui tend à être parallèle aux veines dans
lesquelles il se vide. Ce système lymphatique est particulièrement développé
chez les mammifères.
Les capillaires
lymphatiques convergent au niveau des ganglions renfermant de nombreux types
cellulaires parmi lesquels on retrouve des lymphocytes. Les ganglions
lymphatiques sont le siège de nombreuses réactions immunitaires.
Ø Distribution
des gaz respiratoires :
Compte–tenu de
la faible solubilité de l’O2 dans les
liquides, des débits très importants seraient nécessaires pour assurer la
distribution de ce gaz aux cellules utilisatrices enfouies au plus profond de
l’organisme.
Ainsi, chez la majorité des métazoaires
de grande taille, il existe dans le fluide circulant des molécules qui
augmentent la capacité de transport de l’O2. Ce sont des
pigments respiratoires (ou pigments sanguins)
§ Transport du dioxygène :
·
Les
différents types de pigments :
Les
pigments sont des métalloprotéines comme les cytochromes cellulaires impliqués
dans le transport d’électrons ou d’autres enzymes utilisant l’oxygène
moléculaire comme substrat (catalase, peroxydase) Il existe plusieurs types de
métalloprotéines :
-
Les
protéines avec 1 ou plusieurs atomes de métal liant l’oxygène :
-
Les
hémocyanines (2 atomes de Cu2+),
-
Les
hémérythrines (2 atomes de Fe2+)
-
Les
protéines avec un groupement prostétique (= tétrapyrrole) associé à un atome de
fer ferreux (Fe2+) :
-
Les
hémoglobines,
-
Les
cholocruorines.
Ces pigments
peuvent être en solution dans le sang ou encore dissous dans le cytosol.
¨
Hémoglobines :
Ces
protéines présentent une très forte parenté avec les myoglobines dissoutes
(dans les cellules musculaires lisses de vertébrés et d’invertébrés, de même
que chez les champignons, les plantes et de nombreux protistes, voire des
bactéries)
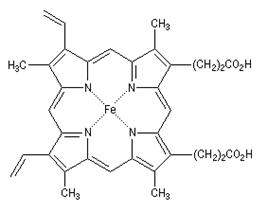
L’hème
se trouve dans une poche hydrophobe de la protéine.
L’hémoglobine
non oxygénée a une couleur rouge sombre. L’oxygène se lie de manière
réversible. L’hémoglobine oxygénée (= oxyhémoglobine) a une couleur rouge vif.
Chez
les vertébrés, l’hémoglobine possède 4 sous–unités de type myoglobine (a2b2) Le tétramère
est capable de lier 4 molécules d’O2.
La fixation dans
le tétramère devient coopérative (courbe sigmoïde) L’affinité d’un site libre
augmente lorsque les autres sont liés à une molécule d’O2.
La transition
allostérique est une modification de conformation.
La fixation peut
être modulée par des effecteurs, comme les protons qui baissent l’affinité.
Chez les
invertébrés, il existe des hémoglobines intracellulaires (ressemblant à
l’hémoglobine des vertébrés)
Chez
certains annélides, échinodermes ou mollusques, il existe aussi des
hémoglobines extracellulaires, de très haute masse moléculaire, correspondant à
des assemblages quaternaires de très nombreuses chaînes (jusqu’à 200) avec
jusqu’à 7 types différents de chaînes chez le lombric et l’arénicole.
Chez les vestimenfères
« riftia » (polychètes) vivant à proximité des sources
hydrothermales, l’hémoglobine transporte l’O2 et le S2– (nécessaire au
métabolisme des micro–organismes symbiotiques)
Chez les
mollusques et crustacés, l’hémoglobine est construite selon un modèle différent
de celui des annélides : les unités de myoglobines sont associées (2 à 18
sites de fixation)
¨
Chlorocruorines :
La forme
oxygénée est de couleur verte et la forme désoxygénée est jaunâtre. Elle a une
structure proche de l’hémoglobine mais l’hème est différent.
¨
Hémérythrines :
Elles sont
présentes dans 4 phyla : les annélides (magellona), les siponculiens,
les priapuliens et les brachiopodes.
-
Les
siponculiens sont des espèces vermiformes marins, vivant dans des hèbes.
-
Les
priapuliens sont des vers prédateurs marins annélés, non segmentés.
La forme
oxygénée est de couleur rose et la forme désoxygénée est incolore. Elle est
intracellulaire et est composée d’octamères et de trimères. Chaque chaîne est
reliée à des atomes de Fe2+.
¨
Hémocyanines :
La
forme oxygénée est de couleur bleue et la forme désoxygénée est incolore. Ce
sont des formes solubles avec 2 atomes du Cu2+ par sous–unité.
Elles sont présentes chez les
arthropodes (crustacés), chitons, quelques bivalves (prosobranches) Elles ont
des motifs décamériques (1 et 2)
·
Capacité
de transport de l’O2 et
modulation de cette capacité :
¨
Affinité du
pigment pour l’O2 :
Elle peut être
exprimée par
%
saturation = 100.[HbO2]
/ ([Hb] + [HbO2])
Les hémoglobines
d’invertébrés ont généralement une P50 plus faible que celle des vertébrés. Par
ailleurs, les hémoglobines des vertébrés (carnivores et insectivores) les plus
actifs ont des P50 plutôt élevées.
Cependant, la
capacité de transport d’O2 ne dépend pas
seulement de l’affinité mais également de la concentration en pigment.
¨
Facteurs
affectant la capacité de transport :
Effet de la température :
Si la
température augmente,
Ce paramètre est important pour les
animaux ectothermes : l’augmentation de la température entraîne une
augmentation de l’activité métaboliques et donc une dissociation de l’O2 plus facile au
niveau des tissus.
Effet du CO2 et du pH :
Les
hémoglobines et hémocyanines sont affectées par les variations de pH. Une
diminution du pH entraîne un déplacement des courbes vers la droite. Cela a
pour effet d’entraîner une dissociation de l’O2.
Les variations
du pH provoque un changement des charges des protéines, d’où une modification
de la conformation spatiale du pigment.
L’effet du pH
est plus marqué chez les poissons (effet Root), il est très important dans le
fonctionnement de la vessie natatoire.
Il existe 2 types d’hémoglobine :
une sensible au pH et l’autre non.
La libération de
CO2 par les tissus entraîne ne
acidification locale et favorise la dissociation de l’O2. Au niveau de
l’échangeur respiratoire, l’élimination de CO2 décale
l’équilibre vers la gauche et donc favorise la fixation de l’O2.
Effet du CO2 :
Le CO2 possède
également un effet direct (indépendamment de l’effet sur le pH) à travers la
formation de composés carbaminés NH–COO– avec les
pigments (amine terminale)
Ce n’est pas le cas avec l’hémoglobine
des poissons et des amphibiens qui ont des composés acétylés.
Effet du pH et le
Il s’agit d’une
action facilitatrice du CO2 et du pH dans
la décharge de l’O2 vers les tissus
à partir des pigments circulants.
L’amplitude
de l’effet de Bohr est variable selon l’espèce : DlogP50/DpH est en relation avec la taille
de l’organisme.
L’effet est plus prononcé pour les
animaux de petite taille, ce qui est important pour un approvisionnement accru
en O2 pour ces animaux qui ont un
métabolisme par unité de masse plus élevé.
Effet des ions :
Il peut être
important chez les invertébrés osmoconformes, qui ne régulent par ou peu leurs
concentrations ioniques.
Effet de petites substances organiques :
Certaines
entraînent une augmentation de
-
Pour
les poissons : ATP et GTP,
-
Pour
les oiseaux : inositolpentaphosphate,
-
Pour
les mammifères : 2,3_prostaglandine et biphosphoglycérate.
Cela concerne
les animaux chez lesquels les hémoglobines sont intracellulaires.
Effet de la production des pigments :
-
Il
peut y avoir un changement du type de pigment au cours du développement, lors
du passage du stade larvaire à l’état adulte.
-
Chez
les vertébrés, il peut avoir une augmentation de la synthèse de pigments qui
nécessite l’intervention de l’érythropoïétine (= facteur de croissance naturel
stimulant la fabrication d’hématies) qui favorise l’érythropoïèse.
-
Il
y a aussi une mobilisation des hématies dans la rate.
NB :
L’O2 peut être
stockée dans les tissus : myoglobine, fluides cœlomiques.
§ Transport du CO2 :
Le CO2 est un gaz
soluble. La majorité est combinée à l’eau :
H2O + CO2 D H2CO3 D H+ + HCO3– D 2H+ + CO32–
Lente
Rapide
Dans les
cellules sanguines (érythrocytes et hématies), il y a la présence d’anhydrase
carbonique qui accélère l’hydratation du CO2.
Au niveau des tissus, les protons H+ se combinent
avec l’hémoglobine, formant du HHbO2 qui exerce un
effet de Bohr entraînant la décharge d’O2. Le CO2 est également
fixé sous forme carbaminée.
Au niveau des poumons et des branchies, il y a les
réactions inverses : il y a la diffusion du CO2 vers l’air et
l’O2 entre dans le sang. L’anhydrase
carbonique convertit les HCO3– du plasma en H2O et CO2 dans les
cellules endothéliales et les hématies.
Régulation :
Chez
les vertébrés à respiration aérienne et chez les insectes, le taux de CO2 prédomine dans
le contrôle de la respiration.
Chez les
vertébrés et invertébrés aquatiques,
-
La
concentration en O2 est plus faible
dans l’eau que dans l’air.
-
L’O2 est moins soluble
dans l’eau que le CO2, il n’y a pas
de problème d’évacuation du CO2.
v Introduction –
généralités :
La
locomotion est l’une des caractéristiques les plus frappantes des animaux. Le
mouvement de l’organisme dans son entier est bien souvent une condition pour se
procurer de la nourriture et permettre la rencontre des partenaires sexuels
pour assurer la reproduction.
Les espèces ont développé des modalités
particulières pour attirer la nourriture (courant d’eau, association
symbiotique, parasitisme) et pour la reproduction l’émission de gamètes dans le
milieu ambiant (espèces aquatiques)
La
locomotion d’un animal dérive d’une poussée contre le milieu ambiant (ou le
substrat) qui, compte tenu de la 3ème loi de Newton, génère une
force égale et opposée contre l’animal.
Une partie de cette force permet le
mouvement (principe de l’action réaction) Il y a transfert de force entre
l’animal et son environnement.
Le mouvement est restreint par :
-
L’inertie :
C’est la tendance d’un objet à demeurer
dans un état statique s’il est stationnaire ou, au contraire, à demeurer en
mouvement s’il bouge. L’inertie est proportionnelle à la masse du corps et
concerne essentiellement les animaux de grande taille.
-
Les forces de
frottement liées à la viscosité du milieu :
La viscosité
exprime la tendance des molécules à adhérer ensemble. La viscosité d’un milieu
est un paramètre qui affecte davantage les animaux de petite taille.
Les
forces de résistance qui s’appliquent à un objet en mouvement dépendent et sont
proportionnelles à :
-
La
vitesse,
-
La
viscosité du milieu,
-
La
surface de l’objet,
-
Et
sa forme.
D’où la
nécessité de forme hydrodynamique ou aérodynamique pour les animaux de grande
taille pour lesquels le rapport surface/volume est faible.
-
Résistance faible à la
pression -
Surface importante Þ
fortes forces de friction
Haute
pression Basse pression
-
Résistance élevée à la
pression -
Faible surface Þ
faibles forces de friction
(turbulences)
Il n’est donc
pas surprenant que les petits animaux planctoniques ne soient pas
hydrodynamiques.
Pour
se déplacer dans les milieux fluides (eau, air), les animaux rencontrent des
problèmes liés à l’absence de substrat solide qui supporte leur masse. Par
conséquent, la densité (et la viscosité) du milieu a une incidence sur la
locomotion au niveau de son coût énergétique.
Chez les animaux aquatiques de grande
taille, il y a eu le développement de stratégies pour diminuer leur densité
(flottabilité)
Dans
le cas des déplacements terrestres, le transfert de force dépend de la réaction
du substrat (déformation, coefficient de frottement)
Le mouvement dépend des caractéristiques
du milieu, mais aussi du plan d’organisation de l’animal et de sa taille.
Chez
les métazoaires, la poussée initiale est généralement générée par :
-
Le
battement de cils,
-
Des
changements de forme,
-
Des
ondulations du corps,
-
Le
mouvement d’appendices.
Pour
les 3 derniers cas, la force est produite par la contraction de muscles. Pour
permettre le déplacement, ces derniers doivent s’insérer sur un squelette
rigide résistant à la déformation :
-
Squelette
osseux des vertébrés,
-
Exosquelette
des arthropodes / coquille des mollusques,
-
Hydrosquelette
des annélides, etc.
Hydrosquelette =
compartiment à parois souples rempli de liquide corporel (incompressible)
Les parois sont
formées de muscles reliés par du tissu conjonctif. La contraction des muscles
met le liquide sous–pression : le compartiment redevient rigide. Si une
partie de la paroi e un résistance moindre, il y a allongement du mouvement.
Pour les protozoaires :
Ils utilisent
des mouvements amœboïdes (amibes), et des mouvements ciliaires (ciliés,
flagellés)
Pour les métazoaires :
|
Déplacements dans
un milieu liquide |
Nage sans
appendices squelettiques (squelette
hydrostatique) |
-
Nage à réaction (méduses, céphalopodes) -
Ondulations (annélides) -
Nageoires souples (gastéropodes) |
|
Nage avec
appendices squelettiques |
-
Battements d’appendices (arthropodes) -
Battements d’appendices et ondulations (poissons,
vertébrés) |
|
|
Déplacements à
la surface du substrat |
Reptation |
Gastéropodes,
reptiles (serpents) |
|
Marche |
Arthropodes,
vertébrés |
|
|
Déplacements
dans l’air |
Vol, saut |
Arthropodes,
vertébrés |
v Bases moléculaires du
mouvement :
Le mouvement
cellulaire implique les éléments constitutifs du cytosquelette (microtubules,
microfilaments) Le cytosquelette est responsable de la forme mais également des
mouvements intracellulaires (chromosomes, organites) ou des mouvements de la
cellule par rapport à son milieu.
Ø Microfilaments :
-
Microfilaments
fins (actine),
-
Microfilaments
intermédiaires (kératine, vimentine, desmine, etc.),
-
Microfilaments
épais (myosine)
L’interaction
entre les filaments d’actine et de myosine est à la base de la contraction
musculaire, d’autres acteurs moléculaires sont requis : la tropomyosine,
la troponine, le Ca2+ et l’ATP.
L’actine et la myosine sont protéines contractiles qui génèrent une force. La
tropomyosine et la troponine régulent l’interaction déclenchée par la
libération de Ca2+ entre l’actine
et la myosine. L’hydrolyse de l’ATP fournit l’énergie nécessaire à la
relaxation.

NB :
Les muscles
striés (que l’on trouve, par exemple chez les vertébrés, dans les parois des
vaisseaux, l’iris, les muscles excréteurs des poils, ou les viscères) ne sont
pas impliqués dans les mouvements du corps et présentent des myofilaments
« dispersés » et non alignés.
Les
fibres musculaires du muscle squelettique des vertébrés se divisent en 2
catégories distinctes :
-
5µm : fibres dites « rapides » ou
résistantes, contenant peu de myoglobine (peu colorées)
" Mouvements
rapides.
-
10 – 20µm : fibres dites
« lentes » ou endurantes, riches en myoglobine (rouges)
" Force de
contraction peu importante lors d’exercices longs.
Un
muscle peut être constitué de l’une ou l’autre de ces fibres ou les 2 à la
fois.
Le
mouvement amiboïde (= amœboïde) est une caractéristique de certains
protozoaires, sarcodines, rhizopodes (= foraminifères et amibes) De même, on
retrouve cette propriété dans les cellules cœlomiques et sanguines de nombreux
métazoaires. Ce phénomène s’observe également dans les tissus embryonnaires.
Le mouvement amiboïde met en jeu une
transition gel D sol réversible
du réseau d’actine de la cellule (= changement de viscosité de la cellule)

-
A
l’avant de la cellule, la profiline accélère la polymérisation de l’actine
augmentant la viscosité (gel)
-
A
l’arrière de la cellule, la cofiline induit la dépolymérisation réduisant la
viscosité (solution)
D’autres protéines
(a_actinine
et filamine) organisent l’actine en réseau gélatineux. La myosine lie ce réseau
à la membrane plamsique.
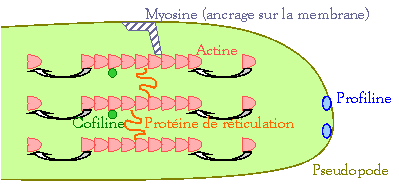
SOL GEL
Ø Microtubules :
Ils sont
présents dans le cytoplasme de toutes les cellules eucaryotes. Ils proviennent
de la polymérisation de protéines globulaires : les tubulines a et b. Ils se forment
par allongement de microtubules « germe » (= a–tubuline)
pré–existant au niveau des centres organisateurs des microtubules (= COMT) à
proximité des centrioles, à la base des cils et des flagelles.
Structures impliquées :
-
La
forme des cellules (axopodes des héliozoaires),
-
Les
mouvements intracellulaires (organites, chormosomes),
-
Les
constituants permanents des cellules (centrioles mais aussi des cils et des
flagelles)
Les
flagelles sont impliqués dans la locomotion des cellules isolées ou des petits
métazoaires, ou encore dans le cas de cellules épithéliales dans les courants
de fluides (eau, mucus, etc.)
Les cils et les
flagelles ont la même structure :
-
Les
cils sont généralement nombreux et présentent un axonème court (région
protubérante)
" Mouvement
rapide dans un sens et lent dans l’autre (rôle de déplacement du milieu par
rapport à la cellule)
-
Les
flagelles sont peu nombreux et à axonème long (par rapport à la taille de la
cellule)
" Mouvements
ondulants (rôle de déplacement de la cellule par rapport au milieu)
L’interaction
entre les bras de dynéine du microtubule a avec le
microtubule b
adjacent, couplée à l’hydrolyse de l’ATP par la dynéine permet le glissement
des doublets périphériques et donc une courbure de l’axonème (à travers une
succession de couplage/découplage)
Le mouvement est lié au battement de
l’axonème.
Ø Aspects
énergétiques :
Les interactions
entre les molécules du cytosquelette nécessitent de l’énergie fournie
essentiellement par l’hydrolyse de l’ATP. Dans le cas de la fibre musculaire
striée, il existe plusieurs voies métaboliques permettant la régénération de
l’ATP à partir de l’ADP.
§ Voie directe par
transphosphorylation :
Myokinase
(adénylkinase)
![]() 2 ADP ATP
+ AMP
2 ADP ATP
+ AMP
§ Transphosphorylation à partir
de phosphagènes :
Phosphagène = molécule avec une liaison
phosphate riche en énergie
-
Créatine–phosphate
chez les vertébrés,
-
Créatine–phosphate
ou arginine–phosphate ou autre phosphagène chez les invertébrés.
![]() Phosphocréatine
kinase
Phosphocréatine
kinase
ADP +
phosphocréatine ATP
+ créatine
Phosphoarginine
kinase
![]() ADP +
phosphoarginine ATP
+ arginine
ADP +
phosphoarginine ATP
+ arginine
La
transphosphorylation à partir de phosphagènes constitue une source immédiate de
phosphates à haute énergie.
La répartition de créatine–phosphate et
d’arginine–phosphate est aléatoire chez les invertébrés et n’est pas liée à la
phylogénie.
 Bien
qu’abondants, ces phosphagènes sont très vite consommés et dans le cas d’un
effort musculaire soutenu, l’énergie provient nécessairement du catabolisme du
glycogène et des acides gras.
Bien
qu’abondants, ces phosphagènes sont très vite consommés et dans le cas d’un
effort musculaire soutenu, l’énergie provient nécessairement du catabolisme du
glycogène et des acides gras.
v Mouvements en milieu
liquide (nage) :
Ø Nage
à l’aide de cils :
Ce
type de locomotion ne concerne que les animaux de petite taille présentant un
grand rapport surface/volume. On la retrouve donc chez les protozoaires ciliés
ou flagellés, mais également chez les métazoaires ciliés de petite taille
comme :
-
Les
rotifères,
-
Les
gastrotriches,
-
Les
petits annélides,
-
Les
larves d’invertébrés (mollusques, échinodermes, larves trocophore des annélides)
Ce
type de déplacement ne s’observe jamais chez les arthropodes et les nématodes
(pour ces 1ers, on a longtemps pensé qu’ils ne possédaient pas de
cils fonctionnels, mais on a observé que les yeux possèdent des cils modifiés)
Notons que ce mode de déplacement concerne
également les spermatozoïdes d’un grand nombre d’animaux qui nagent au moyen de
flagelles.
§ Ciliature uniforme :
La
ciliature forme des vagues métachronales (= qui se déplacent le long de
l’animal)
C’est
le cas de la paramécie.
Les
larves peuvent présenter une ciliature uniforme. Il s’agit des larves
lécitotrophes chargées de vitellus :
-
La
larve miracidium des trématodes (douves),
-
La
larve planula des cnidaires,
-
La
larve parenchymula des éponges (rondes)
§ Ciliature en bande ou en anneau :
C’est le cas de
la larve trocophore (en forme de toupie) des annélides et en 1er
stade larvaire chez les mollusques présentant un développement larvaire.
§ Ciliature sur les lobes ou
les bras :
Ciliature sur les lobes :
C’est le cas de
la larve planctonotrophe des mollusques bivalves et des gastéropodes à
développement urinaire (au 2ème stade = stade véligère) dont les
cils sont présents au niveau du velum (organe locomoteur servant aussi à la
nutrition)
La
ciliature permet un mouvement vertical en spirale. Les cils s’agglomèrent en
cirres avec l’évolution de la larve et permettent un mouvement plus régulier.
Cependant, la croissance de la coquille
tend à limiter la propulsion de la larve qui se transforme en larve
pédivéligère (caractérisée par la présence d’un pied) et abandonne la vie
pélagique pour se fixer. La fixation de la larve déclenche alors la
métamorphose.
Ciliature sur les bras :
Il
y a la présence de bras ciliés servant à la locomotion et à la nutrition :
-
La
larve bipinnaria (astérides),
-
La
larve ophioplutéus (ophiorides),
-
La
larve echinoplutéus (crinoïdes)
Ø Nage
à réaction :
Exemples :
§
La méduse :
Chez les
diploblastiques, il n’existe pas de muscle à proprement parler. Cependant, les
cellules épithéliales possèdent des fibres musculaires au niveau basal (en
contact avec la mésoglée) Il n’existe pas de trop gros problème de flottabilité
pour ces animaux riches en eau.
§
La coquille St
Jacques :
Elle effectue un
battement brusque des valves par contraction du muscle adducteur. La relaxation
lente des muscles permet l’entrée d’eau.
§
Les céphalopodes
(calmar et seiche) :
Les nageoires
postérieures ou latérales servent de stabilisateurs pour la nage rapide, les
tentacules servent de gouvernail. L’ondulation des nageoires assure la nage à
faible vitesse. La cavité palléale peut se remplir d’eau. L’eau est chassée par
le siphon suite à la contraction du manteau.
Cas
du calmar :
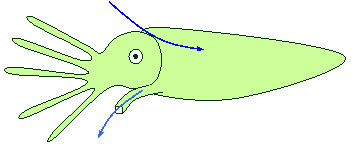 Le manteau n’est
pas fixé à la tête du coté ventral. L’eau pénètre dans la cavité palléale par
des fentes.
Le manteau n’est
pas fixé à la tête du coté ventral. L’eau pénètre dans la cavité palléale par
des fentes.
L’entonnoir peut
être orienté si bien que la nage par réaction peut se faire en marche avant ou
arrière. En réaction de fuite, la vitesse peut atteindre 40km/h, soit la
vitesse d’un saumon de même taille. Le manteau constitue un squelette
hydrostatique.
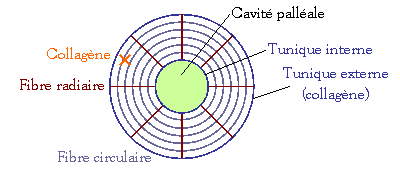
La
contraction des muscles augmente la pression dans la cavité palléale. Il existe
des fibres circulaires à contraction lente pour assurer une nage lente. Les
fibres circulaires centrales et radiales sont rapides pour une nage rapide
(fuite)
Le calmar présente une structure
« ballastes » contenant une solution de NH4Cl.
NB :
Si un animal
nage avec une densité plus élevée que celle de l’eau, il dépense une partie de
son énergie pour ne pas couler et une autre pour son déplacement. S’il peut
avoir une flottabilité neutre (même densité que l’eau), il dépense alors moins
d’énergie pour son déplacement.
Cas
de la seiche :
Près
de 10% de son volume corporel est occupé par la coquille interne « os de
seiche » dont la densité est proche de 0,6. Il présente une structure
laminaire :
-
Des
fines lamelles de CaCO3 maintenues par
des piliers et renforcées par de la chitine,
-
L’espace
entre les lamelles est rempli de gaz (N2 et O2) permettant une
même pression que dans les liquides corporels. Il résiste à des pressions
proche de 25atm (260m de profondeur)
L’eau n’entre
pas dans la coquille grâce à un mécanisme de transport actif d’ions qui assure
une pression osmotique capable de compenser la pression hydrostatique et la
pression osmotique du milieu.
Le
transport actif en ions Na+ et Cl– permet une
faible pression osmotique à l’intérieur par rapport à l’extérieur. Si la
différence de PO (DPO) est
supérieure à la pression hydrostatique, alors l’eau sort et il y a la diffusion
des gaz contenus dans les liquides corporels. Si l’eau est pure, la seiche est
capable de s’opposer à une pression hydrostatique de 25atm (260m)
Cela permet une grande liberté de
mouvement à la verticale sans avoir à ajuster le mécanisme de flottaison.
Ø Nage
par ondulation :
C’est
le cas des animaux dont le corps est allongé. Les ondes de contraction sont
engendrées par la contraction des muscles striés sur l’axe squelettique (corde,
cœlome ou squelette hydrostatique)
Ces mêmes mouvements d’ondulation
provoquent la reptation en milieu solide. C’est le cas des nématodes, serpents,
anguilles, annélides (oligochètes et hirudinés)
Ø Nage
à l’aide de nageoire souple :
Ce type de nage
est observé chez les animaux à corps mou. Il est répandu chez les mollusques
gastéropodes opistobranches ptéropodes. Ils possèdent des expansions latérales
du pied (parapodies) comme chez le « papillon de mer ». C’est le cas
également des opistobranches (aplysie ou lièvre de mer)
L’ondulation des
parapodies est assurée par une augmentation de la pression de l’hémolymphe (mouvements
lents)
Ø Nage
par battements d’appendices :
C’est le cas en
partie des crustacés nageurs comme les crevettes par battements de leurs
appendices abdominaux : les pléopodes. Ce sont des appendices aplatis,
allongés et recouverts de soies.
Plein (grec) = nage
Chez
les larves de crustacés, les antennes, les mandibules ou les appendices
thoraciques sont utilisés pour la nage. Les larves ne possèdent pas tous les
appendices de l’adulte.
C’est le cas également des crabes
portunidés qui nagent grâce à leur 5ème paire de pattes thoraciques
en forme de pagaie.
Les insectes
d’eau douce sont également bons nageurs comme le dytique (coléoptère) dont la 3ème
paire de pattes aplaties munies de soies sert de rame.
Chez
les vertébrés, on observe aussi une nage par battements d’appendices pour les
semi–aquatiques. Exemples :
-
Le
canard avec ses pattes palmées,
-
Les
tortues dont les appendices pectoraux sont palmés,
-
Les
grenouilles avec leurs longues jambes, leurs pieds palmés et la peau recouverte
de mucus (diminuant la friction)
Ø Nage
par ondulation et battements d’appendices :
C’est le cas de
la plupart des vertébrés bons nageurs.

Les
nageoires paires sont à faible implantation (oscillantes) ; les nageoires
simples sont à large implantation (ondulantes) La nageoire caudale joue le rôle
de gouvernail.
Les poissons ont une forme
hydrodynamique. Ils sont recouverts de mucus permettant une diminution de la
friction, une protection contre les ectoparasites et une diminution des
échanges d’eau avec le milieu (étanchéité)
Les
muscles du tronc sont des myomères segmentés dont la contraction provoque des
ondulations latérales du tronc et de la queue. Ces ondulations concernent
surtout la partie postérieure du tronc et la nageoire caudale pour les espèces
dont le corps est court, ou encore pour les nageurs rapides (comme le thon) Les
ondulations concernent également les nageoires à grande implantation.
Les myomères des poissons sont
constitués de muscles lents (rouges) pour le rythme de « croisière »
et de muscles rapides (blancs) pour les accélérations et échappements.
Les
nageoires médianes, de même que les nageoires paires, assurent une stabilité
contre le roulis et empêchent aussi la partie antérieure du poisson de bouger
de gauche à droite (à cause de l’ondulation)
Les oscillations des nageoires paires et
l’ondulation des nageoires médianes permettent les manœuvres.
Un
poisson doit être capable de rester à une profondeur particulière et également
de maintenir sa stabilité. Le foie des requins (par exemple) contient des
quantités considérables d’huile (scalène = C30H70 = précurseur du
cholestérol) qui peut représenter 75% de la masse de cet organe.
Dans la mesure où le scalène a une
densité d’environ 0,86 et comme, par ailleurs, le quelette est cartilagineux et
donc léger (absence de Ca3(PO4)2), la
conjonction de ces 2 éléments assure une flottabilité pratiquement neutre. Un
déficit de portance est assuré par les nageoires pectorales, l’aplatissement
ventral de la tête (effet hydroplanning) et l’asymétrie de la nageoire caudale.
Bon nombre
d’ostréichtiens et de téléostéens possèdent une vessie natatoire remplie de gaz
qui permet d’assurer une flottabilité neutre. La vessie natatoire est un sac à
parois souples situé dans la cavité abdominale sous la colonne vertébréale. Le
volume occupé représente environ 5% (pour les espèces marines) à 7% (pour les
espèces dulcicoles) du volume corporel.
Problème :
La flottabilité
est neutre, c’est–à–dire en équilibre, à une profondeur donnée.
-
Si
le poisson nage en dessous : il y a une compression du gaz et une
augmentation de la densité. Le poisson doit nager activement pour éviter de
couler d’avantage.
-
Si
le poisson nage au dessus du niveau neutre de flottaison : la pression
hydrostatique diminue, la vessie se dilate et la poussée vers le haut augmente.
Certains
poissons ont une vessie natatoire reliée à l’œsophage, ce qui permet qu gaz de
s’échapper. Pour ces poissons, dits physostomes, le remplissage peut se faire
en surface en avalant de l’air. D’un point de vue embryonnaire, la vessie est
une évagination du tube digestif.
Pour d’autres
poissons, dits physoclites, la communication disparaît. Cela concerne
essentiellement les poissons présentant une flottabilité neutre en profondeur.
Les gaz proviennent du sang et sont sécrétés dans la vessie à une pression
égale à celle qui existe à la profondeur où vit le poisson.
Kleistos (grec) = fermé
Comment se fait
la sécrétion de gaz et la rétention de ce gaz dans la vessie natatoire ?
La vessie :
PTOT = 50atm
P(O2) = 40atm
P(N2) = 10atm
Eau – Sang
artériel :
P(O2) = 0,2atm
P(N2) = 0,2atm
+ PHYDROSTAT = 51atm
Þ Ce qui
correspond à un enrichissement en gaz de 200 fois en O2 et de 12,5 fois
en N2.
L’irrigation
de la vessie par le sang artériel devrait conduire à une diffusion des gaz de
la vessie vers le sang. Une structure nommée rète mirabile constitue un piège
pour retenir les gaz par un système d’échange à contre courant (= glande à gaz
= amplificateur à contre courant)
Avant d’atteindre la vessie natatoire,
l’artère se divise en un très grand nombre de capillaires parallèles. Les
capillaires qui sont issus de la glande sont également ramifiés et disséminés
au milieu des capillaires artériels qui se réunissent en un seul vaisseau.
Pour
qu’il y ait diffusion de gaz dans la vessie, il faut que la pression partielle
dans le sang soit supérieure à la pression partielle dans la vessie. La glande
sécrète de l’acide lactique, faisant baisser le pH, diminuant l’affinité de
l’hémoglobine pour l’O2 (effet Root ou
effet de Bohr), ce qui permet d’augmenter
L’O2 diffuse donc
dans les capillaires artériels, comme la vitesse de fixation de l’O2 est lente (10 à
20 sec) par rapport à la vitesse de dissociation. A l’équilibre,
De plus, l’acidité provoque le phénomène
de dégazage.
NB :
Certains
poissons benthiques ne possèdent pas de vessie natatoire.
Chez
les autres vertébrés :
-
Les
crocodiles (reptiles/diapsidés/archosauriens/crocodiliens) présentent une queue
compressée latéralement dont le battement sert à la nage. Les appendices
servent de stabilisateurs.
-
Les
mammifères marins présentent une hyperphalangie des membres antérieurs.
v Mouvements à la surface du
substrat (substrat solide) :
" Milieu
terrestre ou aquatique (fond de mer ou lac)
Ø La
reptation :
On
la rencontre chez les animaux dépourvus d’appendices. Le mouvement s’appuie sur
l’hydrosquelette.
-
Chez
les animaux à corps allongé, le mouvement se fait par l’intermédiaire d’ondes
musculaires (annélides, acœlomates et les serpents)
-
Chez
les gastéropodes (escargots et limaces), le mouvement est assuré par
l’ondulation de la partie aplatie du pied : la sole pédieuse.
-
Chez
les animaux terrestres, en particulier, il existe une friction importante au
niveau du substrat. Par conséquent, la production de mucus (polysaccharides)
sert de lubrifiant et protège également l’organisme.
Le mucus est
aussi un élément important pour lutter contre la dessiccation, ce qui est important
pour le maintien du squelette hydrostatique. Le mucus provoque des adhérences.
Chez les gastéropodes, ces propriétés mécaniques peuvent varier rapidement de
l’état solide (accrochage) à l’état liquide (glisse)
Chez
les petits gastéropodes :
-
La
progression peut se faire à l’aide de cils qui battent dans le mucus et l’eau
(surtout chez les espèces aquatiques)
-
La
progression peut se faire à l’aide de muscles et de cils (comme chez la limnée)
-
La
progression peut se faire à l’aide de muscles (comme chez l’escargot helix)
Il
existe 2 types d’ondes :
-
Les
ondes directes : dans le même sens que le déplacement (chez la majorité
des gastéropodes)
-
Les
ondes rétrogrades : dans le sens inverse du déplacement (chez les
gastéropodes prosobranches comme la patelle et chez les polyplacophores comme
le chiton)
La
contraction des muscles dorso–ventraux provoque la liquéfaction du mucus qui se
solidifie à l’arrivé de l’onde (relâchement des muscles) La cavité hémocœlienne
se trouve comprimée par les muscles : les lacunes sanguines closes
représentent l’hydrosquelette.
Les muscles
transverses empêchent une déformation vers le haut et assurent aussi une
rigidité et permettent ainsi un mouvement longitudinal.
C’est un principe similaire pour les
ondes directes.
Les ondes
peuvent concerner toute la surface du pied ou de manière alternée d’un coté de
l’autre du pied.
Cas d’annélide terrestre :
le ver de terre
Dans la mesure
où les vers de terre vivent dans le sol, le déplacement s’apparente au
fouissage. Le métamérisme des annélides est une adaptation pour le déplacement
(reptation) et l’enfouissement par déformation du corps. Le liquide cœlomique
constitue l’hydrosquelette.
Chez le ver de
terre, la contraction des muscles augmente la pression du fluide cœlomique,
entraînant la rigidification du corps, ce qui provoque l’élongation par
l’étirement des muscles longitudinaux. La contraction des muscles longitudinaux
augmente la pression mais provoque le raccourcissement du corps.
La
compartimentation du cœlome du ver de terre et d’autres annélides permet le
confinement de ce processus (allongement de quelques segments) Des ondes
d’élongation (contractions préristaltiques) se déplacent vers l’arrière de
l’animal.
Mode particulier de déplacement
différent de la reptation :
Certaines
sangsues nagent, d’autres sont retournées hors de l’eau (dans les forêts
tropicales) et se déplacent grâce à leur ventouse. Certaines chenilles se
déplacent de la même manière.
Ø La
marche :
Cela concerne
les animaux avec des appendices (pattes) Elle correspond à l’action de faire
passer le poids d’un corps d’un pied à l’autre. La course effective d’une patte
alterne avec celle de l’autre patte.
Animaux à pied mou avec un squelette
hydrostatique :
C’est
le cas des échinodermes. Ils se déplacent à l’aide de pieds ambulacraires (ou
podia) :
(1) Contraction du
muscle entraînant un flux de liquide vers le pied qui s’allonge (mais qui de
s’élargie pas compte tenu de la présence de collagène),
(2)
Contraction
des muscles rétracteurs entraînant le raccourcissement et la courbure du pied.
Animaux à squelette rigide :
-
Chez
les myriapodes comme le mille–pattes, il y a des mouvements coordonnés des
appendices (ondes métachronales)
-
Chez
les arachnides (4 paires), 2 paires sont au sol et les 2 autres en l’air.
-
Chez
les décapodes comme les crabes (5 paires dont une parfois hypertrophiée), il y
a des mouvements rapides. Toutes les pattes ne sont pas utilisées.
-
Chez
les insectes (3 paires), il y a un appui en trépied.
-
Chez
les vertébrés, la majorité est tétrapode. Les amphibiens, reptiliens et oiseaux
n’utilisent que les appendices postérieurs (bipédie) Les mammifères (sauf
l’homme) utilisent un appui 3 pattes pour avancer la 4ème.
Ø Substrat
liquide :
Cela concerne
les animaux légers dont les extrémités des pattes possèdent des soies
hydrofuges permettant la glisse sur l’eau, comme certains insectes qui se
déplacent à la surface de mares calmes.
Autre cas : les siphonophores qui
sont des hydrozoaires possédant des flotteurs.
v Locomotion en substrat
meuble : fouissage :
Ø Animaux
à corps mou : utilisation du squelette hydrostatique :
Exemple d’acœlomates :
Les anémones de
mer utilisent sa cavité gastrique comme un squelette hydrostatique (contraction
du sphincter) pour s’enfouir dans le sable.
Cœlomates :
Ver de terre,
mollusques bivalves lamellibranches (coques, couteau), scaphopodes (dentale)
Phases
successives de fouissement chez un bivalve (sable ou vase) :
(1)
Pénétration
par ouverture de la coquille (relâchement des muscles adducteurs)
Les valves prennent appui sur le sable et
s’enfoncent. Le pied est poussé vers l’avant grâce à la contraction des muscles
transverses.
(2)
Les
muscles adducteurs se contractent, les valves se ferment et chassent l’eau de
la cavité palléale.
La contraction des muscles protracteurs
augmente la pression dans l’hémocœlome qui entraîne un gonflement du pied et
crée ainsi un 2nd appui.
(3)
La
contraction des muscles rétracteurs du pied va tirer l’animal.
Ø Animaux
à corps rigide :
Les
arthropodes et les vertébrés utilisent des appendices squelettiques.
-
Excavation
du trou,
-
Dégagement
du matériel excavé,
-
Nécessité
de placer les appendices non impliqués hors du champ d’excavation.
Les appendices
sont utilisés comme des pelles et sont souvent aplatis et élargis.
Exemples :
-
Les
crustacés malacostracés amphipèdes ne possèdent pas de carapace. Leur corps est
compressé latéralement.
-
Les
puces des sables et de mer utilisent les appendices thoraciques 2 et 3 (=
gnathopodes) Le 2nd gnathopode est élargi en forme de pelle et les
pléopodes chassent le sable.
-
Chez
certains chélicérates, les araignées peuvent creuser à l’aide des chélicères
qui présentent un sillon et une surface dentée.
-
Certains
insectes s’enfouissent également comme le « criquet taupe ».
Chez la courtilière, la 1ère
paire de pattes est large en forme de pelle dont les tibias et les tarses sont
adaptés pour couper les racines dont ils se nourrissent.
Les termites
(isoptères) creusent le bois qu’ils ingèrent et forent des tunnels
interconnectés dans lesquels ils vivent.
Chez les vertébrés :
Ce
mode de locomotion est retrouvé chez :
-
Les
amphibiens anoures (crapauds et grenouilles),
-
Les
reptiles (tortues),
-
Les
mammifères (taupes, hérissons, fourmiliers, tatous, rats, rats taupes,
blaireau, etc.)
La
forme du corps tend à être cylindrique. Les incisives ont une croissance
continue chez les rongeurs. Ils présentent des griffes et des phalanges
spatulées.
Les mammifères fouisseurs présentent des
membres courts et puissants avec des mains et des pieds proportionnels grands.
-
La
taupe dorée creuse grâce à son museau transformé en pelle.
-
Le
campagnol mord et arrache son substrat.
-
Les
amphibiens anoures creusent en bêchant avec leurs membres postérieurs
(glyphoglossus)
-
Le
fourmilier crochète la terre dans une infractuosité du substrat grâce aux
griffes les plus développées des membres antérieurs.
Le
fouissage permet aussi :
-
Un
refuge diurne pour les formes nocturnes,
-
Un
refuge nocturne pour les formes diurnes,
-
Un
refuge saisonnier pour les espèces hibernatrices,
-
Un
garde manger.
Il
ne devient un mode de locomotion que pour les vertébrés qui mènent en
permanence une existence souterraine et qui sont confrontées à des conditions
de survie bien définies :
-
Faible
pression de prédation,
-
Ressources
relativement abondantes,
-
Obscurité,
-
Forte
hydrométrie,
-
Réflexes
mécaniques liés au déplacement.