
Retour au sommaire des cours |

Une version pdf est à votre disposition ici. |
Embryologie : �tude
du d�veloppement d�un organisme depuis l��uf f�cond� (= cellule �uf) jusqu��
l��tat adulte, sans oublier le processus de vieillissement.
La fonction de reproduction :
caract�re qui d�finit le vivant �
p�rennit� des esp�ces
����������� Sexu�e ou asexu�e.
Reproduction asexu�e :
les individus sont g�n�tiquement semblables aux parents sous r�serve de
l�incidence des mutations.
����������� Elle est observ�e chez les
eucaryotes simples (algues unicellulaires et Protozoaires)
����������������������� = reproduction par mitose.
Elles est plus
g�n�ralement r�pandue chez les animaux ayant une organisation fonctionnelle
simple comme les M�tazoaires Diploblastiques (�ponges)
et certains Triploblastiques
(�chinodermes, Ann�lides)
Certaines
esp�ces pr�sentent une reproduction alternant phase sexu�e et asexu�e.
v Reproduction
sexu�e :
Phase de f�condation :
ph�nom�ne de fusion de 2 gam�tes haplo�des de m�me esp�ce.
����������� �
formation d�une cellule �uf = zygote (diplo�de)
Formation de l��uf :
=
aboutissement des gam�togen�ses ♂ et ♀
=
d�but de construction d�un nouvel organisme
�
maintien de l�esp�ce (ce n�est pas une fonction vitale pour l�individu)
Elle est
observ�e chez presque tous les M�tazoaires.
L�individu
fils re�oit un g�nome nouveau (la moiti� de chacun de ses parents)
� recombinaison g�n�tique.
La
reproduction de l�individu est exclusivement d�volue aux gam�tes (= cellules
sp�cialis�es uniquement dans cette fonction)
En g�n�ral,
les gam�tes ♂
et ♀
sont produits par des individus distincts : esp�ce gonochorique.
����������� ����������� ♂ :
testicule �
spermatozo�des
����������������������� ♀ :
ovaire �
ovule
� esp�ce hermaphrodite (Mollusque,
Vers) :
����������������������� ♂♀
: ovotestis �
spermatozo�des et ovules
En g�n�ral, la
f�condation est crois�e (� autof�condation)
Les 2 sexes
produisent des cellules germinales primordiales (C.G.P) = pr�curseurs des
gam�tes.
![]()

![]() Individu
mature����� ����������� C.G.P������������������������������������������������������������ ����� Spermatogen�se
Individu
mature����� ����������� C.G.P������������������������������������������������������������ ����� Spermatogen�se
Sexuellement������������������������������������ M�iose = r�duction
chromatique (R.C)������ �Ovogen�se


Cycle haplo-diplobiontique������������������������������
Gam�tes
� Croissance
����������� Cons�quences : ��������� Modification des combinaisons
g�n�tiques,
���������������������������������������������� Zygote
= assortiment g�nique unique sur des milliards d�autres.
Le patrimoine
h�r�ditaire est reproduit par la cellule �uf. Si une mutation se pr�sente, elle
se perp�tuera de g�n�ration en g�n�ration.
Cela
entra�ne la variabilit� et l�adaptation des esp�ces.
Conditions environnementales (=
�pig�n�tiques) : se surajoutent au patrimoine
h�r�ditaire et se rendent �galement source de variabilit�.
����������������������� �
�volution
L�embryologie est-elle r�ellement
indispensable ?
On la retrouve
dans des applications biologiques diverses :
-
L��tude des m�canismes
mol�culaires et cellulaires lors du passage d�un �tre unicellulaire (cellule
�uf) � un �tre pluricellulaire organis� (= embryon puis adulte)
-
L��tude de l�expression des g�nes
du d�veloppement embryonnaire, g�nes intervenant pr�cocement dans le
d�veloppement embryonnaire :
-
Etablissement de la polarit�
embryonnaire ant�ro-post�rieure (A/P) et dorso-ventrale (D/V),
-
Edification des lignages, voies
de diff�renciation ou modes de formation d�un tissu, d�un organe.
Existence de 4 �tapes pour que l��uf
devienne embryon
v La
f�condation :
= union d�un spermatozo�de et d�un ovule
La cellule �uf est h�t�rog�ne :
����������� Pr�sence
de r�gions � granuleuses, � color�es, � lourdes, etc.
Elle contient toutes les potentialit�s
du futur individu :
=
cellules m�senchymateuses indiff�renci�es :
-
Adipocytes,
-
Fibroblastes,
-
Ost�oblastes,
-
Chondrocytes,
-
Myocytes.
Anisotropie =
tous les axes de l��uf pas identiques.
����������� Chez
certaines esp�ces : d�s le stade cellule��uf, le plan de sym�trie
bilat�ral est d�j� �tabli (exemple : Amphibien)
v Phase
de segmentation :
= division (ou multiplication)
cellulaire.
�
passage de l��tat unicellulaire � l��tat pluricellulaire.
� Caract�ristiques :
Pas d�augmentation de la masse par
rapport � la masse initiale de la cellule��uf, ni mouvement des parties les
unes par rapport aux autres (= phase non
dynamique)
� diminution de la taille des cellules.
v Gastrulation :
� Caract�ristiques :
-
animaux
triploblastiques (= tridermiques) : mise en place
de 3 feuillets embryonnaires :
-
ectoderme, superficiel (peau),
-
m�soderme, interm�diaire
(muscles),
-
endoderme, interne (tube
digestif)
-
animaux
diploblastiques (= didermiques) :
-
ectoderme,
-
endoderme.
-
Mis en place du tube digestif et
de l�architecture de l�organisme (migration cellulaire)
� phase dynamique
A la fin de la gastrulation :
mise en place du syst�me nerveux central
����������� = Neurulation
v Organogen�se,
histogen�se et morphogen�se :
= Mise en place des tissus et organes.
�
processus de diff�renciation cellulaire avec l�expression s�lective des g�nes.
����������� =
une cellule n�exprime que les g�nes n�cessaires � sa fonction biologique.
A la suite de l�embryogen�se :
Une p�riode
fonctionnelle permet l��dification de l�organisme adulte.
Il y a une interaction entre les 3
derni�res phases.
v Caract�ristiques
g�n�rales des �ufs :
Selon animaux :
Oviparit�
absolue : d�veloppement en dehors de l�organisme
mat�riel (P.A.R.O), sans �tablissement de liens histologique.
Viviparit� :
d�veloppement � l�int�rieur de l�organisme maternelle et mise en place de
relation histologiques (placenta)
Divers
degr�s interm�diaires :
-
Ovoviparit� :������� incubation puis �closion (= ponte)
-
Paraviviparit� :���� �closion (= ponte) puis incubation.
-
Formes
structurale et dimensionnelle des �ufs :
Elles sont variables dans le monde
animal.
La grosseur de l��uf d�termine
directement les dimensions de l�organisme qui va se d�velopper.
v Plan
mol�culaire :
�uf = prot�ines, lipides, acides
nucl�iques, glucides.
� Les
prot�ines :
-
de r�serve (pour nutrition de
l�embryon),
�
synth�se par l�ovocyte (origine endog�ne) + origine exog�ne.
-
d�usage (pour le d�veloppement),
-
de structure (enzymes,
pr�curseurs)
� Acides
nucl�iques :
ADN, ARNTOTAUX.
� ARN :
�
ARNR (ribosomaux) :
= 95 % des ARN totaux (les plus
abondants)
La quantit� d�ARNR
est proportionnelle au nombre de ribosomes.
����������� Cellule
normale : 106
ribosomes,
����������� Cellule
� �uf : 107
ribosomes.
�
ARNT (de transfert) :
= 3 % des ARN totaux.
�
ARNNH (nucl�aire
h�t�rog�ne) = ARNM (messager) :
= 2 % des ARN totaux.
et pourtant
ces ARN ont une tr�s importante variation qualitative
� codage d�une dizaine milliers de prot�ines.
= Complexit� de l�information de
l��uf (3,7.107
nucl�otides)
����������� puis
cette complexit� de l�information diminue quand l�individu s�organise.
Les ARNM
codent pour des prot�ines n�cessaires lors de la segmentation :
-
Prot�ines membranaires,
-
Composants du cytosquelette,
+ d�autres ARNM
qui codent des prot�ines impliqu�es dans le m�tabolisme de la r�plication de
l�ADN (= machinerie de traduction)
ADN = ADNNUCLEAIRE
+ ADNCYTOPLASMIQUE
���������������������������������������������������������� =
ADN mitochondriaque (80%)
���������������������������������������������������������� +
ADN baignant dans le cytoplasme (20%)
�
SN ARN :
�
Implication dans les particules d��pissage.
�
ARNINTERFERANT (d�couverte
r�cente) :
= ARNANTISENS
�
emp�che un g�ne de s�exprimer : blocage et d�gradation de l�ARNM
ou de la prot�ine.
���������������������������������� A�������� U�������� G
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() ����������� ARNM
����������� ARNM
ARNANTISENS ![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
���������������������������������� U�������� A�������� C
v Vitellogen�se :
Le vitellus se
situe dans le cytoplasme de l��uf, il sert de r�serve nutritive � l�embryon
durant le d�but du d�veloppement.
Le si�ge de la
vitellogen�se est au niveau de l�ovocyte IAIRE bloqu� au stade
Diplot�ne de la 1�re division m�iotique. On y observe 2
phases :
-
La phase de pr�-vitellogen�se (ou phase de petit accroissement),
(2
� 3 ans)
-
La phase de croissance (ou phase de grand accroissement)
(3
mois)
-
Quand l�ovocyte IAIRE
n�est pas entour� de cellules folliculaires (= nourrici�res) :
P�n�tration de
monom�res (oses, acides amin�s) pour effectuer la synth�se des macromol�cules.
-
Quand l�ovocyte IAIRE
est entour� de cellules folliculaires (= nourrici�res) :
P�n�tration de
substance semi-d�velopp�e (polyoses, polypeptides) dont la synth�se finale
s�effectue dans l��uf.
La composition
chimique du vitellus est variable selon les organismes. On trouve 3
types :
-
le vitellus glucidique,
-
le vitellus lipidique (chez les Poissons),
-
le vitellus prot�ique (la plus
part des �uf)
v Classification
des �ufs :
Qu�il soit
vierge ou f�cond�, la cellule �uf est une grosse cellule sph�rique, d�pourvue
de flagelle. De plus, les r�serves cytoplasmiques sont variables :
-
Le cytoplasme nutritif (r�serves
vitellines),
-
Le cytoplasme formatif (pas de
r�serve nutritive)
L�abondance et
la r�partition du vitellus sont tr�s variables en fonction des esp�ces, ce qui
permet la classification des �ufs.
Ces crit�res
sont tr�s importants pour la d�termination du type de d�veloppement.
-
Vitellus peu abondant, voire
absent :
 ����������������������� Il y a un m�lange des 2
types cytoplasmes.
����������������������� Il y a un m�lange des 2
types cytoplasmes.
La
phase de segmentation implique l��uf dans son enti�ret�, qui va se
compartimenter en cellules de plus en plus petite.
On parle de segmentation
totale et d��uf holoblastique.
-
Vitellus abondant :
Il
a une localisation distincte par rapport au cytoplasme formatif.
![]()
 ����������������������� Le vitellus inhibe les
mitose, la segmentation au niveau du cytoplasme formatif.
����������������������� Le vitellus inhibe les
mitose, la segmentation au niveau du cytoplasme formatif.
���������������������������������� On parle de segmentation partielle ou m�roblastique et
���������������������������������������������� d��uf m�roblastique.
� Les
�ufs holoblastiques :
� Les �ufs al�cithiques (ou al�cithes) :
Chez les Mammif�res
placentaires, la perte du vitellus a �t� secondaire lors de l��volution.
Anc�tre des Mammif�res :
�ufs t�lol�cithes (tr�s riches en vitellus)
� Les �ufs oligol�cithes :
Un peu plus de
vitellus.
Le vitellus
est sous forme de petites granulations qui se r�partissent dans tout le
cytoplasme
�
Chez les �chinodermes (�toile de mer, Oursin)
-
L��uf est polaris� :
-
Le p�le sup�rieur (ou p�le
animal)
=
point d��mission des globules polaires lors de l�ovogen�se.
Pour les �ufs
al�cithes : le point d��mission des globules polaires mobiles.
-
Le p�le inf�rieur (ou p�le
v�g�tatif)
Le noyau est
toujours excentr� vers le p�le animal.
� Les �ufs h�t�rol�cithes :
Vitellus
beaucoup plus abondant.
Le vitellus
est sous forme de plaquettes vitellines qui se r�partissent de mani�re
h�t�rog�ne, mais surtout dans l�h�misph�re v�g�tatif (exemple des Amphibiens)
� Les
�ufs m�roblastiques :
�  Les �ufs t�lol�cithes :
Les �ufs t�lol�cithes :

Le cytoplasme nutritif est distinct du cytoplasme formatif.
= p�le v�g�tatif���������������������������������� = p�le animal (+ noyau)
Exemple :
Poissons et sauropsid�s (= Reptiles et Oiseaux)
� Les �ufs centrol�cithes :
Le vitellus
est centr� et entour� par le cytoplasme formatif.
�
Chez les Arthropodes (Crustac�s, Insectes)
v Les �ufs oligol�cithes : l��uf d�Oursin :
On les observe
chez les �chinodermes des fonds marins (esp�ces gonochoriques)
Ce sont des
�ufs libres en eau de mer, sph�rique, avec un cytoplasme l�g�rement color�.
L��uf est
entour� d�une membrane vitelline mince, transparente, doubl�e ext�rieurement
par une gangue g�latineuse (�
peu de vitellus)
Il pr�sente
une r�partition presque uniforme, l�g�rement orient�e dans l�h�misph�re
v�g�tal.
![]()
![]()
![]()
 Globules
polaires : ils sont �mis dans l�ovaire avant la
ponte.
Globules
polaires : ils sont �mis dans l�ovaire avant la
ponte.
� H.A�������������� La
f�condation est imm�diate apr�s la ponte.
D�s
la f�condation, le pigment jaune orang� de l��uf vierge se concentre et
H.V�������������� forme
l�anneau sous �quatorial.
����������������������� Il y a un antagoniste entre
2 gradients :
-
Gradient des
propri�t�s m�taboliques et des organites
(mitochondries et ribonucl�oprot�ines pour les ribosomes),
-
Gradient vitellin.
v Les
�ufs h�t�rol�cithes :
� L��uf de Grenouille (Amphibiens) :
-
La femelle pond des �ufs vierges
dans l�eau sous la forme de paquet et le m�le �met du sperme������� �
f�condation
-
L��uf est riche en
vitellus : membrane vitelline entour�e d�une gangue g�latineuse externe.
-
Un pigment noir�tre est pr�sent
au niveau du cytoplasme superficiel qui recouvre les ⅔
de l��uf, et est absent dans le dernier ⅓.
=
calotte blanch�tre dont le centre est le p�le v�g�tatif. Elle correspond
� la r�gion la plus dense.
Le centre de
l�h�misph�re pigment� est le p�le animal (reconnaissable par la position des
globules polaires)
-
En position sub�quatoriale (juste
dessus de la calotte blanch�tre), on trouve le croissant gris (ou
d�pigment�) Il permet de reconna�tre la sym�trie lat�rale de l��uf.

![]()
![]()
![]() ������� 30�
������� 30�
-
La f�condation se fait dans le
p�le animal.
-
Les plaquettes vitellines sont
plus concentr�es et plus grandes dans le p�le v�g�tal ; alors qu�elles
sont de plus en plus disparates et de petite taille en se dirigeant vers le
p�le animal.
-
Le gradient de
ribonucl�oprot�ines (R.N.P) est mis en place lors de l�ovogen�se :
�
les ARNT
et les sous unit�s ribosomiques sont tr�s concentr�s autours du noyau de
l�ovocyte.
-
La pr�sence de gradients vitellin
et R.N.P d�termine une anisotropie de l��uf, qui est dit h�t�rog�ne.
-
Avant la f�condation, le noyau de
l�ovocyte est bloqu� en M�taphase II.
-
La f�condation provoque des
modifications avant la segmentation et l��uf acquiert une sym�trie bilat�rale.
-
La p�n�tration du spermatozo�de
d�clenche l��mission du 2�me globule polaire et la fin de la m�iose.
�
L�ovocyte II devient un ovotide, et le noyau de l�ovotide devient le pronucl�us
femelle (ADN d�condens� et nouvelle enveloppe nucl�aire)
-
Le spermatozo�de entre en contact
avec l�ovocyte II au niveau de l�h�misph�re animal :
-
Fusion des membranes plasmiques,
-
Incorporation du noyau
spermatique dans la cytoplasme de l�ovocyte II,
-
D�condensation de la chromatide du
noyau du spermatozo�de,
-
Nouvelle enveloppe.
�
Le spermatozo�de devient le pronucl�us m�le.
-
Le pronucl�us m�le se dirige vers
le pronucl�us femelle (dans l�h�misph�re femelle) gr�ce au syst�me
microtubulaire du spermaster (= centriole proximal), en entra�nant derri�re lui
une tra�n�e spermatique (pigment�e)
-
Fusion des pronucl�i dans
l�h�misph�re animal,
-
Formation d�une membrane de
f�condation (= enveloppe nucl�aire),
-
Rotation d��quilibration (ou
d�orientation) :
�
le p�le animal est orient� vers haut et le p�le v�g�tal vers le bas.
-
Rotation de sym�trisation :
�
apparition du croissant gris sur la surface dorsale du future embryon.
L��uf vierge
pr�sente une sym�trie axiale (axe P.A / P.V) mais pas de sym�trie bilat�rale.
-
L�exp�rience d�une f�condation
artificielle (� l�aide d�une micropipette) montre que, quel que soit l�endroit
du d�p�t du spermatozo�de, le croissant d�pigment� glissera � l�oppos� de la tra�n�e
spermatique.
� L��uf de Dentale (Mollusque) :
La plupart des
Mollusques et des Ann�lides pr�sente une segmentation spirale. L�ensemble des
invert�br�s pr�sentant cette segmentation est appel� Spiralia.
L��uf est
sph�rique et pondu dans le milieu marin. Il poss�de une large ceinture de
granulation rouge brique au niveau de l��quateur (entre l�h�misph�re v�g�tal et
l�h�misph�re animal)
Lors de la
ponte, l�ovocyte est bloqu� en Prophase I (stade diplot�ne), la p�n�tration du
spermatozo�de entra�ne la poursuite de la m�iose.

![]()
![]() Calotte
de cytoplasme
Calotte
de cytoplasme
![]() hyalin
hyalin
v Les
�ufs t�lol�cithes :
� L��uf de Poisson (
L��uf pondu
est un ovocyte II bloqu� en M�taphase II. C�est une sph�re jaun�tre, r�sistante
(protection dans l�eau) et riche en vitellus, sous forme de gouttelettes
lipidiques.
�
hypertrophie au niveau de l�h�misph�re v�g�tal.
Le cytoplasme
formatif, o� se situe le noyau, se pr�sente en forme de disque au niveau du
p�le animal et est appel� disque germinatif
ou blastodisque.

���������������������������������� Vitellus lipidique
� L��uf d�oiseau (
-
Il est riche en vitellus et isol�
du milieu ext�rieur.
Lors de la
ponte, le germe (ou embryon) se d�veloppe.
-
Dans la partie haute de
l�oviducte, l��uf est f�cond� (= jaune) Le vitellus est synth�tis� par le foie
maternelle et est d�pos� en couches concentriques, alternativement jaun�tre
(synth�se diurne �
riche en lipide et en carot�no�des) et blanch�tre (synth�se nocturne)
Chez
-
Le noyau et le cytoplasme
formatif sont en p�riph�rie, vers le p�le animal (= disque germinatif,
blastodisque ou �ventuellement encore appel� cicatricule)
-
Lors du cheminement dans
l�oviducte, l��uf tourne (en g�n�ral) dans le sens des aiguilles d�une montre
et est entour� par plusieurs couches d�albumen (= blanc) Le jaune sera maintenu
dans le blanc par des structures appel�es chalazes.
-
Chalaze situ�e � droite de
l�embryon est dite senestre (enroulement dans le sens inverse des
aiguille)
-
Chalaze situ�e � gauche de
l�embryon est dite dextre (enroulement dans le sens des aiguille)
-
Une partie de l�albumine va �tre
entortill�e en pr�sence d�une condensation mucilagineuse.
-
Au final, on observe 3 membranes
k�ratinis�es :
-
La membrane coquilli�re interne,
-
La membrane coquilli�re externe,
-
La coquille calcaire poreuse
d�pos�e quand l��uf est pondu.
-
La lat�bra :
elle correspond � la masse centrale de vitellus ferm�e initialement.
-
Le noyau de Pander + le
col : chemin de migration de la cicatricule vers la surface du cytoplasme
pendant la croissance de l��uf.
-
Une chambre � air est pr�sente au
niveau du gros bout de l��uf. Elle correspond � un d�doublement des membranes
coquilli�res interne et externe.
-
L��uf est un ensemble clos, mais
les �changes avec l�ext�rieur gazeux et thermiques sont possible.
-
L�embryon a une disposition
perpendiculaire par rapport au grand axe de l��uf avec la t�te orient�e. Le
sens de rotation de l��uf suit le plan dorso-ventral (sym�trie bilat�ral)
-
R�gion ventrale : r�gion au
contact du vitellus.
-
R�gion dorsale : r�gion le
plus � l�ext�rieur (surface du blastodisque)
v Les
�ufs al�cithes de Mammif�res :
-
Lors de la ponte (=ovulation),
l�ovule est lib�r� par rupture du follicule de De Graaf. Il est entour� d�une
membrane pellucide, doubl�e ext�rieurement de
-
Lors de la f�condation, au niveau
de l�amPoule (⅓
externe de l�oviducte), il y a �mission du 2nd globule polaire.
Tous deux sont
�mis dans le p�le animal ; ils sont aussi tr�s volumineux et mobiles sous
la membrane pellucide.
-
L��uf f�cond� migre vers l�ut�rus
par contraction p�ristaltique de l�oviducte et par les mouvements ciliaires de
l��pith�lium tubaire.
v Les �ufs centrol�cithes d�Insectes (
-
L�ovogen�se est r�alis�e, en 8
jours, en collaboration avec des cellules nourrici�res.
-
L�ovaire est constitu�
d�ovarioles m�rio�stiques qui collaborent avec des cellules nourrici�res (en
opposition aux ovarioles pano�stiques qui ne n�cessitent pas de collaboration) C�est
l� que se situent les cellules germinales primordiales.
-
Une cellule germinale primordiale
subit une mitose et donne 2 cellules :
-
Une cellule souche (cellule
germinale primordiale),
-
Une cellule � l�origine de la
chambre ovarienne (ou ovovarique)
-
Apr�s 4 mitoses de la cellule
initiale, on obtient une grappe de 16 cellules � cytodi�r�se (=plasmodi�r�se)
incompl�te. Il y a un contact cytoplasmique entre les 16 cellules, on parle de
ponts cytoplasmiques.
-
La cellule 1, dite sexuelle,
devient le gam�te ♀ qui
grossit et subit la m�iose.
-
Les 15 autres deviennent les
cellules nourrici�res.
-
Les cellules nourrici�res
subissent 8 cycles de r�plication de l�ADN sans mitose. Au d�part, chacune
poss�de 4 lots de n chromosomes (= 2 lots de 2n) � la fin de ces r�plications,
elles poss�dent 1.024 lots de n chromosomes.
On
a donc obtenu 1.024*14 �
15.000 g�nomes identiques qui fonctionnent pour 1 seule cellule et dont le
volume cellulaire est multipli� par 90.000 en 3 jours.
-
Les cellules nourrici�res sont
appel�es cellules polyt�niques (= qui n�ont pas le m�me nombre de chromosomes
que les autres cellules)
-
Elles synth�tiques des ARN et des
prot�ines.
-
Les cellules folliculaires
entourent les cellules nourrici�res et l�ovocyte.
-
Lors de la maturation de
l�ovocyte, il y a une r�sorption des 15 cellules nourrici�res au d�pend de la
cellule sexuelle qui grossit.
-
La chambre ovarienne
= �uf allong�, ellipso�de, riche en vitellus. Elle est recouverte par le
chorion.
-
Le chorion = coque
�paisse et rigide. Il pr�sente un micropyle qui permet le passage du
spermatozo�de contenu dans la spermath�que. Lors de la ponte, il est fix� sur
un support.
v Introduction :
� Caract�ristiques :
-
Il s�agit de la phase qui suit la
f�condation et consiste en une succession de mitoses avec cloisonnement
cytoplasmique. Le germe se compartimente donc en cellules de plus en plus
petites. Ces divisions mitotiques sont appel�es divisions de segmentation ou
clivages.
-
Lors de cette phase, la masse
totale de l�embryon = la masse totale de la cellule �uf.
Lors
de chaque division, la masse d�une cellule fille = � de la masse de la cellule
m�re.
����������������������� �
augmentation du rapport nucl�ocytoplasmique : VNucl�aire / VCytoplasme
-
1er clivage :
2 cellules� filles (= blastom�res) �
partir de la cellule �uf.
-
Le germe passe par le stade
morula (aspect d�une m�re), puis par le stade blastula (apparition d�une cavit�
de segmentation, le blastoc�le)
-
Le type de la segmentation d�pend
de la quantit� de vitellus et donc de la classification des �ufs.
-
�uf t�lol�cithe (vitellus tr�s
important) : p�le inf�rieur de l��uf non segment�.
-
�uf al�cithe, oligol�cithe ou
h�t�rol�cithe : segmentation totale (= holoblastique)
-
�uf t�lol�cithe, ou
centrol�cithe : segmentation partielle (= m�roblastique)
� Segmentation holoblastique :
-
Totale et �gale :
le blastoc�le est entour� par une assise r�guli�re de blastom�res de m�me
taille����������������� ����� �
C�loblastule r�guli�re (exemple de l�Oursin)
-
Totale et in�gale :
le blastoc�le est entour� par 1 ou plusieurs assises de blastom�res de tailles
irr�guli�res.�� ����� �
C�loblastule� irr�guli�re
(exemple des Amphibiens)
-
Blastoc�le virtuel :
il est combl� par des blastom�res v�g�tatifs volumineux
�
Sterroblastule (exemple des Ann�lides)
-
Blastom�res de tailles diff�rentes :
-
![]()
![]() Petites :
microm�res�������������������� s�il y a 2 tailles diff�rentes.
Petites :
microm�res�������������������� s�il y a 2 tailles diff�rentes.
-
Grandes : macrom�res
-
Moyennes : m�som�res��������������� s�il y
a 3 tailles diff�rentes.
Dans le cas
d�une segmentation totale, selon la disposition des blastom�res, on observe une
segmentation soit radiaire, soit spirale.
-
Segmentation totale
radiaire : alternance de clivages m�ridiens
(quartiers de blastom�res verticaux) et latitudinaux (quartiers de blastom�res
horizontaux)
�
sym�trie radiaire = parfaite superposition des
blastom�res les uns par rapport aux autres (exemple de l�Oursin et du d�but de
l��uf d�Amphibien)
-
Segmentation totale
spirale : clivages obliques (chez Spiralia, les Ann�lides
et quelques Mollusques)
� Segmentation m�roblastique :
= segmentation
partielle, soit disco�dale, soit superficielle.
-
Segmentation partielle disco�dale : chez
les �ufs t�lol�cithes (Poissons, sauropsid�s)
Le p�le
inf�rieur n�est pas segment� ; seul le p�le sup�rieur (= disque germinatif
ou embryon) subit des clivages. Les blastom�res sont regroup�s sous forme de
disque appel� blastoderme ou blastodisque.
�
Discoblastule
������� = observation d�une calotte de
blastom�res qui repose sur le vitellus.
-
Segmentation partielle
superficielle : chez les �ufs centrol�cithes (Insectes)
Il y a
multiplication du noyau dans le vitellus sans structure (= syncytium), puis les
noyaux migrent dans le cytoplasme p�riph�rique pour former l�assise syncytiale.
Le blastoderme p�riph�rique entoure le vitellus central qui ne subit pas de
segmentation.
����������� ���������������������������������� �
P�riblastule avec un blastoc�le virtuel
v Segmentation holoblastique des �ufs
oligol�cithes : l��uf d�Oursin :
= Segmentation
totale, �gale et radiaire pour les 3 premiers clivages, puis segmentation
in�gale.
-
1er clivage :
m�ridien (= dans l�axe PA � PV)
� 2
cellules semblables (= blastom�res)
-
2�me clivage :
m�ridien et perpendiculaire au 1er
� 4
blastom�res semblables.
-
3�me clivage :
�quatorial
� 8
blastom�res semblables
(4 dans l�h�misph�re animal, 4 dans l�h�misph�re
v�g�tatif)
-
4�me clivage :
16 blastom�res
-
Pour l�h�misph�re animal : 8
m�som�res dispos�s sur une seule couche.
-
Pour l�h�misph�re v�g�tal :
4 macrom�res (sous l��quateur)
+ 4 microm�res (pr�s du p�le v�g�tatif)
-
6�me clivage :
64 blastom�res
-
Pour l�h�misph�re animal :
16 m�som�res animaux 1 (pr�s du p�le animal)
+
16 m�som�res animaux 2.
-
Pour l�h�misph�re v�g�tal :
8 macrom�res v�g�tatif 1
+ 8 macrom�res v�g�tatif 2
+ 16 microm�res.
�
C�loblastule
(les
blastom�res entourent le blastoc�le qui est mis en place depuis le stade 8
cellules)
La blastula
pr�sente une touffe cili�e apicale (p�le animal) ; le reste du germe est
entour� de cils vibratiles.
v Segmentation
holoblastique des �ufs h�t�rol�cithes :
� Segmentation de l��uf d�Amphibiens :
= Segmentation
totale, �gale et radiaire pour les 2 premiers clivages, puis segmentation
in�gale.
-
1er clivage :
m�ridien (= dans l�axe PA � PV)
� 2
cellules semblables (= blastom�res)
���������������������������������� ����������� Dans 50% des cas, le clivage passe
par le croissant d�pigment�.
Les
blastom�res sont de taille identique mais diff�rents sur le plan mol�culaire et
d�veloppemental (� cause de la pr�sence ou non d�une partie du croissant
d�pigment�)
-
2�me clivage :
m�ridien et perpendiculaire au 1er
� 4
blastom�res.
����������������������� ����������� ��������� Si le 1er
clivage passe dans le croissant, le 2�me n�y passe pas et
inversement.
-
3�me clivage :
latitudinal sus �quatorial
������� (d� � la forte concentration en
vitellus dans l�h�misph�re v�g�tatif)
� 4 microm�res
dans la partie sup�rieure de l�h�misph�re animal,
� 4
macrom�res dans l�h�misph�re v�g�tatif et une partie de l�h�misph�re animal.
-
4�me clivage :
16 blastom�res = stade morula
-
Clivages suivants :
il y a un fort asynchronisme des clivages.
La
segmentation est plus rapide dans l�h�misph�re animal que dans l�h�misph�re
v�g�tatif : les divisions m�ridiennes commencent dans le p�le animal et se
terminent dans le p�le v�g�tatif. Les divisions suivantes recommencent dans le
p�le animal avant que les pr�c�dentes ne soient finies dans le p�le v�g�tatif.
Le vitellus
des macrom�res inhibe les divisions mitotiques.
�
La blastula = plusieurs couches de cellules
(les plus
grosses au niveau du p�le v�g�tatif et les plus petites au niveau du p�le
animal)
De plus, les
macrom�res poss�dent une paroi plus �paisse que celle des microm�res.
-
Au d�but de la segmentation, les
divisions sont extr�mement rapides car il y a absence des phases G1 et G2 dans
le cycle cellulaire. Ce n�est qu�� partir du 10�me au 12�me
clivage qu�il y a r�introduction progressive de ces phases.
-
Le passage � clivages
synchrones � � � clivages asynchrones � est caract�ristique de
la transition blastul�enne (TB ou Mid Blastula Transition) qui
correspond aux 1�res expressions du g�nome de l�embryon.
Lors
de cette transition, les 1�res transcriptions zygotiques coexistent
avec l�ARNM
maternel.
� Segmentation de l��uf de Dentale (Mollusque) :
= Segmentation
totale, in�gale et spirale (plans de clivage obliques, ce qui donne un
arrangement spiral� des blastom�res fils)
-
1er clivage :
presque m�ridien (l�g�rement en biais au niveau du p�le animal)
� 2
blastom�res appel�s� respectivement AB et
CD
(CD est plus gros parce qu�il inclut le lobe
polaire)
-
2�me clivage :
presque m�ridien
AB �
2 blastom�res A et B
CD �
2 blastom�res C et D
(D est plus gros parce qu�il inclut le lobe polaire)
-
3�me clivage :
oblique (45� par rapport � l�axe PA � PV)
� 4
microm�res animaux : 1a, 1b, 1c, 1d.
� 4
macrom�res v�g�tatifs : 1A, 1B,
����������� A �
1a et 1A
����������� B �
1b et 1B������������������������������������ Faisceau
mitotique oblique
����������� C �
1c et
����������� D �
1d et 1D����������������������������������������������� les
cloisons des macrom�res.
Quand on
observe par le p�le animal, les microm�res sont d�cal�s par rapport aux
macrom�res
-
soit dans le sens des aiguilles
d�une montre : spirale dextre,
-
soit dans le sens inverse des
aiguilles d�une montre : spirale s�nestre.
-
4�me clivage :
inversion des faisceaux mitotiques du 3�me clivage �
changement du sens de la spirale.
-
Clivages suivants :
disposition altern�e des blastom�res se maintient : l�obliquit� des
faisceaux est alternativement � droite et � gauche.
�
C�loblastule
-
Lors des 3�me, 4�me,
5�me et 6�me clivages :
4 quartettes de microm�res (et 4 quartettes de macrom�res) sont produites par
les macrom�res.
La division et
la distribution des blastom�res permettent de d�finir une nomenclature
conventionnelle des blastom�res.
Dans la
segmentation spirale, les blastom�res sont dispos�s de fa�on � �tablir une plus
grande surface de contacts par rapport � ce que l�on observe dans la
segmentation radiaire.
En g�n�ral,
les embryons sujets � la segmentation spirale subissent moins de clivages avant
la gastrulation par rapport aux autres modes de segmentation.
v Segmentation
m�roblastique des �ufs t�lol�cithes : l��uf de Poule :
= Segmentation
disco�dale qui d�bute dans la partie haute de l�oviducte.
Les 3 premiers
clivages� sont perpendiculaires � la
surface du cicatricule (= blastodisque) et font une d�coupe en tranches plus ou
moins r�guli�res.
-
1er clivage :
m�ridien qui part du centre du cicatricule vers les extr�mit�s.
-
2�me clivage :
m�ridien, perpendiculaire au 1er plan de clivage
(et perpendiculaire � la surface de l�embryon)
-
3�me clivage :
m�ridien (apparition de 2 sillons lat�raux parall�les au 1er plan de
clivage.
-
Clivages suivants :
latitudinaux, tr�s irr�guliers, et asynchrones.
On obtient des
blastom�res superficiels, individualis�s et s�par�s par une couche (ou assise)
syncytiale. Il y a un contact cytoplasmique entre les blastom�res.
-
Jusqu�au stade 64 cellules,
on se trouve dans le stade morula.
Apr�s, on
arrive dans le stade blastula, caract�ris� par l�apparition d�une cavit� entre
les blastom�res et le vitellus, remplie d�un liquide s�cr�t� par les
blastom�res.
Elle est
appel�e cavit� sous germinale, cavit� de segmentation ou blastoc�le primaire.
-
� la ponte,
on observe une aire pellucide au centre du blastodisque qui correspond aux
blastom�res situ�s au dessus du blastoc�le I AIRE
ainsi qu�une aire opaque qui correspond aux blastom�res dispos�s au dessus, et
au contact, du vitellus.
v Segmentation
m�roblastique des �ufs centrol�cithes d�Insectes : l��uf de
Drosophile :
Dans
l�ooplasme (cytoplasme + vitellus), le noyau de la cellule � �uf subit 8
divisions synchrones rapides. Ces divisions ne sont pas accompagn�es d�un
partage du cytoplasme par des membranes����������� =
processus d�endomitose
�
�formation d�un syncytium
-
Apr�s 8 divisions,
les noyaux commencent � migrer vers la p�riph�rie de l�embryon, le p�riplasme, et les divisions continuent. Les
noyaux sont alors appel�s �nergides.
Lorsque le
d�but de migration s�effectue, on atteint le stade blastoderme syncytien.
-
Apr�s environ 6 divisions
suppl�mentaires, des membranes plasmiques
s�installent et on arrive au stade blastoderme cellularis�. � ce stade, il
y a la 1�re expression des g�nes.
Malgr� tout,
une centaine de noyaux restent localis�s dans le vitellus (central) Ces noyaux
sont � l�origine des vitellophages.
-
Vers le 9�me
clivage : les noyaux sont arriv�s dans la r�gion
post�rieure de l�embryon. Ils sont � l�origine des cellules polaires qui
repr�sentent les pr�curseurs des cellules germinales primordiales. Leur
localisation par rapport � l�embryon est externe. L�embryon est appel�
p�riblastule.
v Segmentation
holoblastique de l��uf al�cithes de Mammif�res :
L�embryon se
d�veloppe lors du trajet dans l�oviducte et aussi lors de l�implantation dans
la muqueuse ut�rine (= nidation)
2
phases : 1 libre et 1 fix�e
La
segmentation intervient lors de la phase libre.
-
1er clivage :
peut � �tre m�ridien. Les globules polaires sont mobiles sous la membrane pellucide ;
or, le p�le animal est d�fini par la position de ces globules polaires.
� 2
blastom�res de tailles diff�rentes : 1 macrom�re et 1 microm�re.
-
2�me clivage et
apr�s : divisions asynchrones. Le macrom�re
fait �a division en 1er.
� 3
blastom�res.������� Puis le microm�re fait
sa division.
� 4
blastom�res : 2 macrom�res et 2 microm�res.
La
segmentation se poursuit lors du trajet de l�oviducte. Les cellules de
-
Stade 8 � 16 cellules :
La morula arrive dans la cavit� ut�rine et elle commence � subir les ph�nom�nes
de cavitation (= formation du blastoc�le) et de compaction (= mise en place de
jonction �tanche) qui permet une perm�abilit� s�lective.����� �
r�le protecteur
-
Les cellules apolaires occupent
le centre de l�embryon et d�rivent des macrom�res.
Les
cellules polaires sont dans la p�riph�rie de l�embryon et d�rivent des
microm�res.
-
D�s la cavitation, l�embryon est au
stade 32 � 64 cellules et devient la blastula (appel� aussi blastocyste)
-
Les blastom�res au centre du
blastocyste sont � l�origine du bouton embryonnaire.
Les
blastom�res p�riph�riques forment le trophoblaste, la paroi embryonnaire qui
est � l�origine du placenta.
L�embryon est
encore libre dans la cavit� ut�rine.
-
Entre le 6�me et le 13�me
jours de nidation : le blastocyste est face � la
muqueuse ut�rine. Le trophoblaste se diff�rencie en syncytiotrophoblaste qui
est en contact avec la muqueuse et en cytotrophoblaste qui est en contact avec
le bouton embryonnaire.
-
Le contact avec la muqueuse est
de plus en plus importante pour que l�embryon �tende ses liens histologiques
avec la circulation sanguine de la m�re (liens directs)
F�condation�� Segmentation����������������������� � Gastrulation������������
Organogen�se
![]()
![]()
![]() ����������� ���
�uf����������������������� Blastula���������������������� Gastrula��������������������� Neurula puis Juv�nile
����������� ���
�uf����������������������� Blastula���������������������� Gastrula��������������������� Neurula puis Juv�nile
Segmentation :
multiplication cellulaire active importante.
Gastrulation :
mouvements morphog�nes pour la mise en place des feuillets embryonnaires
(ectoderme, endoderme et m�soderme pour les triploblastiques)
Organogen�se :
diff�renciation des tissus en des tissus fonctionnels.
v Modalit� de la gastrulation :
� Invagination ou en embolie :
Exemple :
chez les �chinodermes.
= mouvement
d�une partie du blastoderme dans le blastoc�le.
����������������������� �
blaste � : tissu qui est � l�origine de �
� Recouvrement ou �pibolie :
Exemple :
chez les Amphibiens.
=
prolif�ration cellulaire des microm�res du p�le animal = recouvrement des
macrom�res du p�le v�g�tal qui deviennent internes.
� D�lamination :
Exemple :
chez les C�lent�r�s.
=
prolif�ration cellulaire avec un plan de clivage parall�le � la surface de
l��uf.
���������������������������������� Cellules
filles������������������������� ����������� Archent�ron
(ouverture ult�rieure
sur l�ext�rieur)
� Immigration :
Exemple :
chez les Vert�br�s sup�rieurs.
Les cellules
du blastoderme migrent et se rejoignent dans le blastoc�le.


���������������������������������� Archent�ron
(ouverture ult�rieure sur l�ext�rieur)
� Prolif�ration polaire :
Pour les �ufs
t�lol�cithes.
=
prolif�ration cellulaire � l�un des p�les de la blastula.
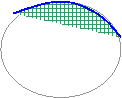
![]() ���������������������������������������������� Blastoderme��� ���������������������������������������������� Ectoblaste
���������������������������������������������� Blastoderme��� ���������������������������������������������� Ectoblaste
![]() ���������������������������������� �� (= disque germinatif)
���������������������������������� �� (= disque germinatif)
��������������������������������������������������������������������������������������������������������������������� Endoblaste
����������� �
Pas d�archent�ron
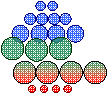
v L��uf
d�Oursin (�chinoderme) :
� Gastrulation :
� la fin de la
segmentation, l��uf est devenu une blastula.

![]()
![]()
![]()
![]()
![]() M�som�res���������������������������������������������������� Cellules
animales 1
M�som�res���������������������������������������������������� Cellules
animales 1
![]()
![]() ���������������������������������������������������������������������� Cellules
animales 2
���������������������������������������������������������������������� Cellules
animales 2
![]()
![]()
![]()
![]() Macrom�res�������������������������������������������������� Cellules v�g�tatives 1
Macrom�res�������������������������������������������������� Cellules v�g�tatives 1
![]()
![]()
![]()
![]()
![]()
![]() ���������������������������������������������������������������������� Cellules
v�g�tatives 2
���������������������������������������������������������������������� Cellules
v�g�tatives 2
![]() Microm�res���������������������������������������������������
Microm�res���������������������������������������������������
![]() Cellules v�g�tatives 2
Cellules v�g�tatives 2
La
gastrulation se d�roule en 3 �tapes :
�
Il y a :
-
Une diminution des divisions
cellulaires,
-
L�apparition d�une touffe
ciliaire apicale au niveau du p�le animal,
-
Un aplatissement et un
�paississement du p�le v�g�tatif formant la plaque v�g�tative,
-
L�immigration des microm�res dans
le blastoc�le formant du m�soblaste (ou m�senchyme primaire)
‚
Il y a :
-
Une embolie des macrom�res
v�g�tatifs 2
�� invagination en doigt de gants formant
l�archent�ron.
-
Un d�tachement des cellules �
l�extr�mit� apicale de l�archent�ron,
�
formation du m�senchyme secondaire qui correspond aux �l�ments sanguins.
ƒ
Il y a :
-
Formation des v�sicules
c�lomiques = ent�roc�lie � partir de v�sicules.
�
mise en place des 3 feuillets = fin de la gastrulation.
� Organogen�se :
= formation de
la larve Plut�us.
-
Le germe (= gastrula) s�aplatit
sur sa face ventrale,
-
La formation de l�anus ventral =
blastopore,
-
Il y a une d�pression stomod�ale
formant le stomodeum (= bouche) par mouvement interne de l�ectoderme.
-
Les bords lat�raux de la bouche
s�allongent en formant 4 bras (2 oraux et 2 anaux)
v L��uf
de Grenouille (les Anoures) :
� Gastrulation :
Rappel :
L��uf subit :
-
Une rotation d��quilibration qui
d�termine le p�le animal (et le p�le v�g�tal)
-
Une rotation de sym�trisation par
rapport au croissant qui d�termine l�axe dorso-ventral.
Apr�s la
segmentation, on obtient :
-
Des microm�res au niveau du p�le
animal,
-
Des macrom�res au niveau du p�le
v�g�tal (riche en vitellus),
-
Et le blastoc�le est d�port� dans
le p�le animal.
� Aspect morphologique in vivo :
-
Au d�but de la gastrulation, il y
a une encoche entre le croissant d�pigment� et le p�le v�g�tatif : la
fente blastoporale (ou blastopore) qui est bord�e dans sa partie sup�rieure par
un bourrelet cellulaire (= l�vre dorsale)
-
La fente blastoporale est due par
une embolie (= mouvement vers l�int�rieur) de quelques blastom�res v�g�tatifs,
suivie par une �pibolie (mouvement de surface vers le blastopore) des cellules
du croissant puis des microm�res de l�h�misph�re animal.
-
Ensuite, la l�vre blastoporale
effectue un mouvement vers le p�le v�g�tatif et va avoir des formes
successives :
-
Anse de panier,
-
Fer � cheval,
-
Circulaire.
-
L�embolie entra�ne la
redistribution des masses cellulaires cr�ant un mouvement de bascule.
-
La fente blastoporale est �
l�origine du future anus.
-
Le croissant d�pigment� et
l�h�misph�re v�g�tatif disparaissent.
-
Il y a la fermeture du
blastopore.
� Mise en place des feuillets et �tude des mouvements morphog�nes :
-
Mouvements internes ?
-
Quelle est la destin�e des
r�gions cellulaires ?
�
Mise en �vidence des
territoires pr�somptifs :
Vogt,
1925 : techniques des marqueurs color�es avec
l�utilisation de diff�rents colorants biologiques� (rouge neutre, sulfate de bleu du Nil)
=
suivi du d�placement des cellules.
-
Actuellement, on utilise un
marquage d�une cellule donn�e (micro-injection) par un colorant
fluorescent� (fluorochrome =
fluoresc�ine) Ensuite, on effectue des coupes histologiques pour conna�tre le
devenir des cellules color�es au cours de la gastrulation.
On
d�finit la carte des territoires pr�somptifs : � tel groupe de
cellules de la blastula donne tel feuillet de la gastrula.
-
Il y a une invagination (=
embolie) de la r�gion inf�rieure : formation du m�soderme et de
l�endoderme par recouvrement total de la gastrula par l�ectoderme.
�
Mouvement de la
gastrulation :
-
� la fin de la
gastrulation : l�ectoderme est superficiel ; le m�soderme et
l�endoderme d�limitent l�archent�ron ; le blastoc�le est enti�rement
envahi par le m�soderme.
-
Ensuite, l�organogen�se d�bute avec
la neurulation (= r�arrangements internes des feuillets m�sodermiques et
endodermiques.
� La neurulation :
= formation de
l�axe nerveux.
� Observations in vivo (= externes) :
(en surface)
En face
dorsale, il y a :
-
un aplatissement de la gastrula,
formant la plaque neurale d�limit�e par des replis (=bourrelets neuraux),
-
un rapprochement de ces
bourrelets lat�raux,
-
une d�pression de la plaque neurale������� �
stade goutti�re neurale,
-
une soudure des bourrelets (=
internalisation de la goutti�re)��� �
stade tube neural,
+
un renflement ant�rieur � l�origine de l�enc�phale,
-
une absence de soudure aux
extr�mit�s formant :
-
un neuropore ant�rieur (fermeture
plus tardive)
-
et un neuropore post�rieur (=
orifice caudal)
� Coupes histologiques :
= observation
des arrangements internes des feuillets.
�
Stade goutti�re
neurale :
�
Stade tube
neural :
M�soderme :
-
S�gr�gation (subdivision) en
:
-
M�soderme para axial,
-
M�soderme interm�diaire,
-
Et lames lat�rales.
-
Cavitation (= formation de
cavit�s) :
-
�pic�lome (dans le m�soderme para
axial),
-
M�soc�lome (dans le m�soderme
interm�diaire),
�
origine du n�phrotome.
-
Hypoc�lome (dans les lames
lat�rales)
Les lames lat�rales se rapprochent sans fusionner et
forment le m�sent�re.
On obtient alors 2 feuillets :
-
Le Splanchnopleure (proche de
l�endoderme)
-
Et le Somatopleure (proche de
l�ectoderme)
-
M�tam�risation (succession
de structures) : formation de somites � partir du
m�soderme para axial.
�
Stade bourgeon caudal :
= formation du
Rhombenc�phale
-
Fermeture post�rieure du tube
neural (il n�y a plus de neuropores),
-
Ouverture de l�anus.
�
Organisation
m�tam�rique apr�s la neurulation :
-
M�tam�risation des cr�tes
neurales (� l�origine des ganglions nerveux),
-
M�tam�risation du m�soderme para
axial (� l�origine des somites),
-
M�tam�risation du m�soderme
lat�ral (� l�origine du n�phrotome)
+
fusion des cavit�s du m�soderme lat�ral pour former une seule cavit� : le
c�lome.
Morphogen�se
des Amphibiens ressemble � la morphogen�se des reptiles, des oiseaux et des
mammif�res.
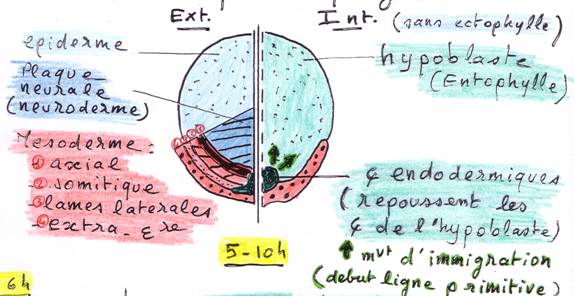
v L��uf
de poule (Gallus domesticus) :
� Pr�gastrulation :
� la fin de la
segmentation, on obtient une blastula primaire.
� la fin de la
pr�gastrulation, on obtient un germe diblastique. Pour cela, il faut 2
migrations :
-
La 1�re migration
forme l�entophylle (ou hypoblaste) primaire,
-
La 2�me migration
forme l�entophylle (ou hypoblaste) secondaire qui correspond au rempart
germinatif sup�rieur.
Cette
pr�gastrulation permet la d�termination des parties ant�rieure (� gauche) et
post�rieure (� droite)
� Gastrulation :
� Observations in vivo (= externes) :
-
5
� 10 H (B) :
����� Formation d�un sillon (= r�gion
�paissie) par des mouvements d��pibolie convergeant vers le sillon et
mouvements d�embolie.
-
16
H (C) :
����������������������� Allongement du
blastoderme dans l�axe c�phalo � caudal.
Allongement
du sillon formant la ligne primitive et une r�gion plus compacte, le n�ud de
Hensen.
-
18
H (D) :
���������������������� Formation du
prolongement c�phalique sous l�ectophyle
(observable par transparence) � l�origine du
m�soderme cordal.
Allongement maximum de la ligne primitive.
-
20
H (C) :
����������������������� Apparition d�un replis
(ou bourrelet) c�phalique ext�rieur,
���������������������������������� Formation
compl�te d�une corde dorsale
(allongement du prolongement c�phalique),
���������������������������������� R�gression de
la ligne primitive
�
positionnement post�rieur du n�ud de Hensen.
La
gastrulation est termin�e. En surface, on trouve le mat�riel ectodermique.
� Mouvements morphog�nes des territoires pr�somptifs :
![]()
![]()
![]()


� Coupes histologiques :
v L��uf
de Drosophile (Insectes) :
� Rappel :
Chez lez �ufs
centrol�cithes, on a :
-
un p�riblaste (= embryon),
-
un ooblaste
-
et un chorion protecteur.
Ils pr�sentent
une segmentation superficielle pour former une p�riblastula.
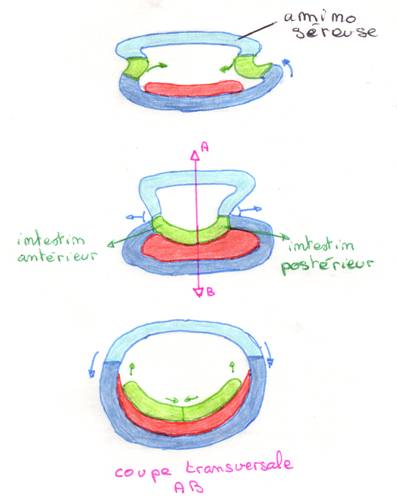
� Gastrulation :
� Les territoires pr�somptifs :
(vue lat�rale
superficielle)
� D�but de la gastrulation :
�
Invagination de l�endoblaste post�rieure.
‚
Invagination du m�soblaste :
����������� �
Formation du sillon ventral m�dian + d�but du recouvrement par
l�ectoblaste.
ƒ
Mise en place du m�soderme et de l�ectoderme embryonnaire :
-
Internalisation du m�soblaste par
le recouvrement de l�ectoblaste.
-
Bourgeonnement et migration des
cellules du m�soblaste
(le recouvrement par l�ectoderme se poursuit)
ƒ
Mise en place de l�endoderme :
-
Invaginations endodermiques
post�rieure et ant�rieure.
v Les
�ufs de Primates (Mammif�res) :
� Pr�gastrulation :
(lors de
l�implantation)
8�me
jour :
-
Apparition de la cavit�
amniotique (formation de l��pith�lium amniotique).
-
Le disque embryonnaire (=
embryon) est didermique :
-
�piblaste = ectophylle
(contact
avec la cavit� amniotique)
-
Hypoblaste = entophylle
(contact
avec le l�cithoc�le primaire)
Du 8�me
au 12�me jour :
-
Apparition de la membrane de
Heuser dont l�origine est le cytotrophoblaste et l�entophylle.
-
Prolif�ration de l�entophylle, le
l�cithoc�le primaire devient alors secondaire et sera � l�origine de la
v�sicule vitelline.
� Gastrulation :
Du 16�me
au 20�me jour ���������� (similaire
aux oiseaux)
� Observations externes :
-
La ligne primitive et le n�ud de
Hensen sont � la surface de l�ectophylle.
-
Les cellules de l�ectophylle
s�invaginent puis s��talent lat�ralement entre l�ectophylle et
endophylle ; elles forment ainsi le m�soderme.
Du 18�me
au 19�me jour : Formation de
la plaque cordale :
-
En r�gion ant�rieure :
fusion du canal cordal avec le toit du l�cithoc�le
(= l�entophylle et fissuration longitudinal de
l�entophylle)
���������������������������������� +
�talement du m�soderme pour former la plaque cordale.
Du 19�me
au 20�me jour : Recul du n�ud
de Hensen et r�gression de la ligne primitive :
�
Allongement ant�ro � post�rieure de la corde (d�tachement des cellules du
m�soblaste de la plaque cordale pour former la corde d�finitive� = un tube plein)
� Neurulation :
Du 19�me
au 21�me jour : diff�renciation du m�soderme et formation de la
goutti�re neurale.