
Retour au sommaire des cours |

Une version pdf est à votre disposition ici. |
Elle est d�finie
comme une enveloppe continue qui forme une barri�re (ou fronti�re) entre le
milieu extracellulaire (= liquide interstitiel) et cytoplasme (= liquide
intracellulaire)
Elle
permet :
-
L�int�grit�
de la cellule,
-
La
r�gulation de la composition du cytoplasme et du liquide interstitiel,
-
Le
maintien des diff�rences de composition chimique.
Exemple : Le sang :
-
Le
liquide interstitiel (= le plasma) est riche en Na+ et pauvre en K+,
-
Le
milieu intracellulaire est pauvre en Na+ et riche en K+.
L�int�rieur de
la cellule est compartiment� en :
-
Milieu
cytosolique,
-
Milieu
interne aux organites,
-
Compartiment
intracellulaire s�par� du cytosol par diff�rentes membranes (membranes
nucl�aire, mitochondriale, du RE, de l�appareil de Golgi).
v Les fonctions de la
membrane plasmique :
� �� :
La membrane est
structure d��changes entre le milieu extracellulaire et le milieu
intracellulaire.
� = Ph�nom�ne de diffusion.
� �‚ :
�
La membrane
r�ceptionne les signaux chimiques r�gulant le fonctionnement de la cellule et
traduit l�arriv�e de ces signaux par l�activation d�enzymes.
� = Ph�nom�ne de transduction.
Exemple : Le mod�le de
transduction du signal lumineux :
![]()
�nergie
lumineuse
![]() ����������������������������������������������������������������� Photor�cepteur (dans la partie profonde de la
r�tine) :
����������������������������������������������������������������� Photor�cepteur (dans la partie profonde de la
r�tine) :
�
l�obscurit� :
������� E = � 30 mV
� D�polarisation
par entr�e de Na+ via des canaux
sp�cifiques ouverts � l�obscurit�.
Cette ouverture
est due � la pr�sence de GMPC (Guanosine Mono
Phosphate cyclique)
� la
lumi�re :
� Changement de
conformation,
������������������ �
Activation de prot�ines G,
������������������ �
Activation d�enzymes effectrices qui vont d�truire les GMPC,
������������������ �
Baisse de la conduction de Na+�� =�� �Diminution du courant d�obscurit�,
� Potentiel de
membrane,
� Influx nerveux.
� �ƒ :
La membrane
permet le maintient de la diff�rence de potentiel �lectrique entre le milieu
extracellulaire et le milieu intracellulaire.
� �„ :
Dans les
cellules excitables, la membrane g�n�re et/ou propage un ph�nom�ne �lectrique.
� �… :
La membrane
plasmique renferme des activit�s enzymatiques qui participent aux voies
m�taboliques de la cellule et � sa r�gulation (= lieu enzymatique)
Exemple : La synth�se de
microfibrilles de cellulose par la Cellulose�synth�tase chez les cellules
v�g�tales.
� �† :
Les composants
de la membrane peuvent servir de substrats � la formation de mol�cules
n�cessaires � la cellule.
Exemple : L�acide
arachidonique des phospholipides membranaires est le pr�curseur de la
Prostaglandine. Les phospholipides sont des acides gras d�une vingtaine de carbones
� doubles liaisons.
� �‡ :
Sur la face
externe de la membrane, se trouvent des composants qui forment des d�terminants
antigoniques dans la reconnaissance tissulaire (implication dans le syst�me
immunitaire)
v Architecture g�n�rale des
membranes biologiques :
Les membranes
sont compos�es de lipides, de prot�ines et de glucides. Le rapport
prot�ines/lipides est variable selon le type cellulaire et l�esp�ce.
Exemple : La membrane
interne des mitochondries ����� � 76 % de
prot�ines.
La
membrane de my�line� � que 18 % de
prot�ines� (isolement �lectrique de la
cellule nerveuse)
Le pourcentage
de phospholipides est non n�gligeable : en quantit� majoritaire (surtout
des glyc�rophospholipides)
� Les
lipides membranaires :
� Les
glyc�rophospholipides :
Ils constituent
40 % du poids total des membranes. On y trouve principalement :
-
Des
glyc�rophospholipides (55%),
-
Du
cholest�rol (20%), il est abondant dans les membranes plasmiques mais rare dans
les membranes intracellulaires,
-
Des
glycolipides (5%),
-
D�autres
lipides dont les sphingolipides (20%)
Les lipides les
plus nombreux et les plus repr�sent�s sont construits � partir de squelettes
d�un mono�ester du glyc�rol. Un nom d�usage leur est attribu� en fonction de
leur 1�re caract�risation (= lieu de leur d�couverte)
Les
glyc�rophospholipides sont pr�sents chez les animaux, les plantes et
micro-organismes :
![]() ������������������ L�cithines,
������������������ L�cithines,
������������������ C�phalines,
������������������ Anositines,
������������������ Etc.
�
Les
glyc�rophospholipides modifi�s :
�
Les
lypoglyc�rophospholipides :
= Agents
h�molytiques et cytolytiques������ � Destruction en
s�incluant dans la membrane.
= Interm�diaires
du catabolisme ou du m�tabolisme des glyc�rophospholipides.
�
Les d�riv�s
�ther � oxydes :
-
Le plasmalog�ne : on le trouve
dans les tissus � haute intensit� respiratoire, les macrophages et les cellules
de la Thyro�de.
-
Le PAF (= Platelit Activating Factor) : c�est le
m�diateur produit par les leucocytes pour activer les plaquettes.
�
Les
propri�t�s physiques des glyc�rophospholipides :
Il s�agit d�une
substance amphiphile : pr�sence d�une t�te polaire, ionis�e, et de 2
queues apolaires. Cela leur conf�re une double affinit�.
Ils sont
solubles dans des solvants organiques, comme le chloroforme et le m�thanol, et
sont insolubles dans l�ac�tone.
Ce sont des
mol�cules transio�actives :
-
Ils
ont une propri�t� cruciale au niveau pulmonaire, � la surface des alv�oles,
dans les �changes gazeux,
-
Et
ils emp�chent le collapsus (= recroquevillement des alv�oles quand elles se
vident)
� Les sphingolipides :
Le squelettes de
ces lipides est form� d�une diolamine, � cha�ne longue de type skingo�de, avec
un acide gras qui se fixe sur le groupe amine (= c�ramide = pr�curseur des
lipides de v�g�taux)
Leur
classification est bas�e sur la nature du groupe R2 li� �
l�hydroxyde :
����������������������������������������� H�������� �������� C�ramides,
����������������������������������������� Pi�������� �������� C�ramides_1_Pi,
������������������������������ Pi_Choline���� �������� Sphingomi�line,
������������������������������ ��� Glucide������ �������� Glycosphingolipides,
����������������������������������������� Ose����� �������� C�r�brosides.
Les sphingosides
et les c�ramides :
Ils sont
construits sur une base sphingo�de.
������ D�riv�s de la sphingosine les plus
importants :
-
Sphingosine
(dans les cellules animales),
-
4_hydroxysphigosine
(dans les cellules v�g�tales),
-
C�ramide
= Sphingoside N_ac�tyl�,
-
Sphingomy�line :
r�le dans la transduction.
�
Les
glycosphingolipides :
-
Lactoglyc�ramide
(cellules c�r�brales),
-
Glycosylc�ramide
(autres cellules),
-
Oligoglycosylc�ramide
(h�maties)
� Les st�ro�des :
= classe
importante dans les lipides membranaires.
On retrouve
surtout le cholest�rol. Il s�agit d�une mol�cule amphiphile qui est � la
fois :
-
Constituant
de la membrane en s�y intercalant et en g�rant sa fluidit�
-
Et,
� l��tat libre dans le cytosol, pr�curseur de mol�cules biologiques comme les
hormones st�ro�des ou les vitamines.
Il est
transport� avec les autres lipides sous forme de lipoprot�ines.
� Les glycoprot�ines :
Il s�agit d�une
association d�un sucre (en g�n�ral un galactose) avec un acide gras au PM �lev�
(la sphingosine)
�
Les
galactolipides :
= 70% du contenu
en lipide de membrane du chloroplaste.
= mol�cule
n�cessaire � la synth�se du syst�me nerveux.
L��nergie
n�cessaire � cette synth�se provient du glucose et du galactose form�s par
l�activit� enzymatique.
�
Les
gangliosides :
Il s�agit de
glycolipides acides dont la cha�ne oligosaccharidique est termin�e par des
structures d�acides sialiques.
Ils sont les
constituants des cellules du syst�me nerveux.
Nomenclature :
|
G |
M , D ou T |
1 chiffre |
|
� Ganglioside � |
1, 2 ou 3
r�sidus d�acide(s) sialique(s) dans la cha�ne oligosaccharidique |
Nombre de
r�sidu(s) d�ose(s) |
�
Les
glycolipides neutres :
= C�r�bro�des.
Ils sont form�s
d�une longue cha�ne et un ose li� par une liaison osidique � la fonction alcool
IAIRE.
-
Galactoc�r�brosides (avec
un b
� galactose) : dans le cerveau,
-
Glucoc�r�brosides (avec
un b
� glucose) : dans les autres organes.
Ils sont
synth�tis�s � partir de sucres activ�s par des glucosyltransf�rases.
� La
bicouche phospholipidique :
= unit�
structurale de base des membranes biologiques.
= 2 couches de
mol�cules phospholipides, constitu�es :
-
D�une
t�te polaire regardant vers le milieu aqueux
-
Et
d�une cha�ne d�acide gras constituant un domaine hydrophobe.
Chaque couche
est appel�e feuillet.
Elles sont
structur�es par :
-
Des
interactions hydrophobes entre les cha�nes,
-
Des
interactions de Van der Waals entre les atomes pour un compactage des parties
hydrophobes,
-
Des
liaisons hydrog�nes et des interactions �lectrostatiques� entre les groupes de t�tes polaires et la
mol�cule d�eau pour la stabilisation de la bicouche.
Dans un milieu
aqueux, les lipides membranaires forment des lysosomes.
La bicouche
poss�de la fluidit� bidimensionnelle d�pendant de sa composition et de la
temp�rature.
Dans la
membrane, il y a une agitation thermique des mol�cules par :
-
Rotation
libre des phospholipides et glycolipides autour de leur axe,
-
Diffusion
lat�rale dans le feuillet = translation.
L�orientation
axiale est � l�origine de l�anisotropie membranaire (= propri�t�s physiques des
phospholipides diff�rant dans toute la structure de l�espace)
Les mol�cules
phospholipidiques en phase lamellaire sont anim�s de divers mouvements et
chacun de ces mouvements est caract�ris� par ses temps de
corr�lation (temps d�effectuer p)
-
Diffusion
rotationnelle,
-
Diffusion
lat�rale,
-
Diffusion
transversale = flip � flop (plus lente car il faut que la t�te polaire doit
traverser la partie hydrophobe)
La membrane est
anim�e dans son ensemble par un mouvement dit � collectif �, appel�
ondulation de surface.
Le lysosome est
une mol�cule sph�rique � compartiment aqueux central et � bicouche
phospholipidique.
� Effets de la
temp�rature :
�
Changement brutal des propri�t�s physiques.
Hausse
de temp�rature : Transition de phase par une mobilit� accrue des
liaisons C�C des cha�nes d�acides gras.
�tat
de gel �
�tat fluide plus d�sorganis�.
Baisse
de temp�rature : �� ����������� plus difficile � supporter pour la
cellule qu�une hausse.
Les cellules animales
et bact�riennes s�adaptent � une baisse de temp�rature du milieu en augmentant
le rapport acides gras insatur�s / acides gras satur�s de la membrane.
Le cholest�rol
membranaire est un facteur d�terminant du maintient de la fluidit� de la
bicouche en s�ins�rant entre les mol�cules de lipides. Il est trop hydrophobe
pour former � lui seul la structure en feuillet. Il est constitu� :
-
D�une
partie hydrophobe polaire en contact avec la phase aqueuse
-
Et
une partie polycyclique qui interf�re avec la structure phospholipidique et
tend � l�immobiliser.
� Les
prot�ines et les glycoprot�ines membranaires :
= 20 � 30% de la
masse membranaire, selon les tissus et les esp�ces.
� Les fonctions des prot�ines
membranaires :
-
Fonction enzymatique :�� plus de 40 activit�s r�pertori�es et
identifi�es.
� Transduction
des messages hormonaux ou nerveux.
-
Fonction de transport membranaire de substance :�����
Elle sert de
canaux sp�cifiques en ayant des fonctions de transporteurs.
-
Fonction capable de liaisons � des mol�culaires informatives :
= r�cepteurs.
-
Fonction capable de liaisons structurales���������� en unissant le cytosquelette � la
membrane.
Exemple : filament
d�actine.
-
Fonction capable de fixer des virus ou des
toxines :
�
Absorption de virus sp�cifiques par glycoprot�ines.
Exemple : le virus SIDA
effectue une liaison au lymphocyte T4 par
l�interm�diaire de la prot�ines SD4.
� Les prot�ines
intrins�ques :
Ou int�gr�es.
Elles forment
des structures ayant une ou plusieurs parties ins�r�es dans la bicouche
(domaines transmembranaires), une partie d�passant du cot� externe et une autre
du cot� inverse.
On peut les extraire apr�s un traitement physique ou chimique qui d�truit la structure membranaire.
Elles poss�dent :
-
Des parties hydrophobes, pour s�associer
aux lipides et permettre une meilleure coh�sion de la membrane, constitu�es
d�aminoacides hydrophobes (Alanine, Isoleucine, Leucine, Proline et Valine)
-
Des r�gions hydrophiles, en contact avec
les milieux interstitiel et/ou cytosolique, constitu�es d�aminoacides
hydrophiles (Arginine, Aspartate, Glutamate, Histidine, Lysine, S�rine, etc.)
-
Et
d�un maximum de liaison entre les groupes C=O et N�H des
acides amin�s constitutifs qui ont un caract�re polaire�������������� � doubles h�lices.
� Zones +/�
hydrophiles.
Elles pr�sentent
le type structural des canaux transporteurs : leur structure est impliqu�e
dans le processus de communication chimique.
� Les prot�ines
extrins�ques :
Ou p�riph�riques
(� l�extr�mit� de la membrane)
Elles sont
constitu�es de domaines essentiellement en contact avec le milieu polaire
(aqueux) La plus part de ces prot�ines sont +/� repli�e sur elles�m�me avec les
cha�nes les plus hydrophiles � l�ext�rieur.
Elles sont
souvent associ�es � un segment hydrophobe permettant leur insertion dans la
membrane par une liaison covalente avec un acide gras, un sucre ou un r�sidu
isoprop�nyle.
� Les
glucides :
= 5 � 10 % du PM
de la membrane.
Ils sont
accroch�s � la membrane par 2 moyens :
-
Fixation
� la surface des prot�ines
-
Ou
fixation aux lipides.
� Leur r�le :
Reconnaissance : � Les motifs glycosidiques sont tr�s
antig�niques.
Exemple : groupes
sanguins.
Participation �
l�environnement local ��� (mol�culaires
tr�s polaires)
Renforcement de
la membrane : ����������������������� Formation
d�un feutrage.
Exemple : � La cellulose solidifie la membrane.
v La membrane plasmique, un
mod�le de la mosa�que fluide :
Singer et
Nicholson, 1972.
La base de la
structure membranaire = bicouche contenant :
-
Des
r�gions hydrophobes (� l�int�rieur)
-
Et
des r�gions hydrophiles (� l�ext�rieur)
" Structure de
lipides fluides : mol�cules entre les 2 interfaces qui maintiennent la
structure en feuillet.
Cette structure
est rigidifi�e par des mol�cules de cholest�rol qui forment un squelette.
� Mosa�que � :
parce que les prot�ines membranaires sont nombreuses et vari�es.
Chez les Eucaryotes,
la distribution des ions de part et d�autre de la membrane est in�gale.
Exemple :
|
|
Int�rieur de
la cellule |
|
Ext�rieur de
la cellule |
|
K+ |
[ K+ ]I |
> |
[ K+ ]E |
|
Na+ |
[ Na+ ]I |
< |
[ Na+ ]E |
|
Ca2+ |
[ Ca2+ ]I |
< |
[ Ca2+ ]E |
La diff�rence de gradient de concentration est due � un ph�nom�ne de transport passif par diffusion.
v La diffusion simple passive
de petite mol�cules � travers une membrane :
= Un solut�
franchit la bicouche lipidique sans interaction sp�cifique avec un compos�
membranaire (il faut que le solut� soit une substance liposoluble)
� La
loi de Fick :
Elle est �tablie
pour tout transfert � travers de la membrane, d�un tissu.
Le d�bit de
transfert est proportionnel � la surface et � la diff�rence de pression (ou de
concentration) et inversement proportionnel � l��paisseur.
La vitesse de ce
ph�nom�ne est proportionnelle � une constante de diffusion qui d�pend (pour le
passage membranaire) de la liposolubilit� et de la taille de la mol�cule.
Capacit� de
diffusion :����� v� =� D . S
. dc
/ e
������ Avec :� D : Coefficient de solubilit� (dans notre cas,
liposolubilit�),
������������������ S � : Surface d��change,
������������������ dc :
Diff�rence de concentration de part et d�autre de la membrane,
e � : �paisseur.
Exemple :�� au niveau pulmonaire :
La quantit� d�O2 qui entre dans
une cellule sera donc plus grande :
-
Si
la cellule est grande,
-
Si
la membrane est mince,
-
S�il
existe une forte diff�rence de concentration de part et d�autres de la
membrane.
Le flux
unidirectionnel est mesur� au moyen d�isotopes radioactifs.
On peut d�duire
la loi de Fick :
En absence de diff�rence de
concentration, les flux unidirectionnels sont �gaux : il n�y a pas de flux
net.
� Le flux diffusionnel :
Il est toujours
proportionnel � la concentration en solut�, quelque soit la concentration de
celui�ci. On n�observe jamais de cin�tique de saturation.
![]()
 v
v
����������������������������������������������������������������� Pour
une substance dont le PM < 500 Da
![]()
����������������������������������������������������� dc
Le flux
diffusionnel peut varier avec chaque substance et pour une substance avec
chaque membrane.
Le caract�re
plus ou moins liposoluble des solut�s et leur plus ou moins grand diam�tre
(encombrement st�rique) vont intervenir pour d�terminer la facilit� plus ou
moins importante � traverser la membrane.
La perm�abilit�
des solut�s d�int�r�t biologique dans les bicouches est faible voire nulle. Les
bicouches sont imperm�ables :
-
Aux
ions inorganiques,
-
Aux
mol�cules polaires charg�es,
-
Aux
acides amin�s (charg�s),
-
Aux
grosses mol�cules polaires non charg�es (glucose, saccharose),
-
Et
aux nucl�otides (encombrement st�rique)
� Passage par la phase
huileuse et pores :
Il y a une
relation �troite entre la perm�abilit� � diff�rentes substances et leur
coefficient de passage huile � eau (= indice de liposolubilit�)
����������������������������������������������������� � Solubilit� du solut� dans la membrane
![]() Coefficient de
partage� =� a� =
Coefficient de
partage� =� a� =
����������������������������������������������������� Solubilit�
du solut� dans le milieu aqueux
� Notion de pores :
Les petites
mol�cules ayant un certain caract�re de solubilit� dans l�eau (hydrophiles)
passent plus rapidement la membrane ; pourtant elles ont une
liposolubilit� faible�
Travaux de
Collander, 1930 :
Le passage se
fait autrement que par la phase huileuse : la notion de pores est un
r�seau de mailles assurant une certaines continuit� de la phase aqueuse entre
les milieux intracellulaire et extracellulaire.
D Les pores ne sont pas des canaux ioniques ! Ils
sont diff�rents des porines (= structure en cha�ne b) Par exemple : les aquaporines assurent le transport de l�eau.
� Les principaux facteurs de
la diffusion simple :
La surface
d��change :
Certaines
cellules sp�cialis�es dans les �changes (cellules intestinales, r�nales)
pr�sentent des caract�res morphologiques pour augmenter la surface d��change
(microvillosit�s, invaginations)
Le poids
mol�culaire :
La membrane
plasmique est pratiquement imperm�able aux substances au PM sup�rieur � 1000
Da.
L�ionisation :
La double couche
lipidique est extr�mement imperm�able aux mol�cules charg�es.
La liposolubilit� :
Il s�agit du
facteur le plus d�terminant.
Exemple :�� La m�thadone = l�apia�de
synchitique :
Elle a une grande
liposolubilit� : le d�but d�action est rapide et la dur�e d�action est
longue.
���� "�� il y a une distribution rapide dans les
tissus.
L��paisseur de
la membrane :
Elle est presque
constante pour toutes les membranes.
v Exemple
de diffusion passive � travers une membrane : l�Osmose :
1830 : mise
en �vidence par le Dr Dutrochet, m�decin naturaliste.
L�osmose a un
r�le important dans les m�canismes physiologiques de la cellule au cot� des
transferts actifs.
� D�finition :
= Transfert du
solvant d�une solution dilu�e vers une solution concentr�e au travers d�une
membrane dite � perm�able �. Dans les organismes vivants, des
transferts d�eau par osmose s�effectuent � travers de la membrane des cellules.

������������������ A������������������������������������������� B�������������������������������������������� C
A :� Un tube est submerg� � moiti� dans
l�eau : l�eau est au m�me niveau � l�int�rieur qu�� l�ext�rieur du tube.
B :� L�extr�mit� du tube est scell�e par une
membrane semi�perm�able, le tube est � moiti� rempli d�une solution sal�e et
submerg� initialement au niveau de la solution sal�e.
C :� L�eau monte dans le tube.
" La membrane est
capable de permettre le passage de l�eau mais pas du sel.
" La hausse de
niveau est attribu�e � la pression osmotique. L�osmose cr�e progressivement l��galit�
de concentration des 2 cot�s de la membrane semi�perm�able.
Dans les
organismes, toutes les cellules jouent le r�le de membrane semi�perm�able par
rapport au liquide dans lequel elles baignent. Le passage de l�eau et des
�l�ments qui l�accompagnent (ions NA+, K+, Mg2+) entre
l�int�rieur et l�ext�rieur est r�gul� par le ph�nom�ne d�osmose.
Ce dernier est
utilis� par les techniques de dialyse r�nale, permettant d��liminer les toxines
(ur�e, acides uriques) et l�eau contenues en trop grande quantit� dans le sang.
� La
pression osmotique :
= Pression qu�il
faudrait exercer sur une solution pour emp�cher le solvant de diffuser au
travers d�une paroi semi�perm�able qui s�pare la solution et le solvant pur.
La pression
osmotique P a une expression similaire � celle de la pression donn�e par
la Loi des Gaz parfaits.
� Cons�quences de l�osmose
pour la cellule :
L�eau diffusant
librement au travers de la plus part des membranes, une cellule peut alors �tre
plac�e dans un milieu isotonique, hypotonique ou hypertonique.
|
Milieu : |
D�finition : |
Isotonique |
Milieu de m�me
pression osmotique que le milieu intracellulaire. |
|
Hypotonique |
Milieu dont la
pression osmotique est plus faible que la pression intracellulaire = La
concentration totale en solut� est plus faible dans le milieu extracellulaire
par rapport au milieu intracellulaire |
|
Hypertonique |
Milieu dont la
pression osmotique est plus forte que la pression intracellulaire = La
concentration totale en solut� est plus �lev�e dans le milieu extracellulaire
par rapport au milieu intracellulaire |
Les cons�quences
sont un �quilibre hydrique dans les cellules :
|
Milieu : |
Cons�quences : |
Isotonique |
Pas de
mouvement net d�eau au travers de la membrane plasmique "� Le volume cellulaire reste stable. |
|
Hypotonique |
Il y a une
entr�e d�eau dans la cellule =� Turgescence |
|
Hypertonique |
Il y a une
sortie d�eau dans la cellule =� Plasmolyse |
Cas des h�maties :
La turgescence
entra�ne l�h�molyse : l�h�moglobine se r�pand dans le milieu
extracellulaire formant des � ghosts � (= globules fant�mes)
d�apparence translucides.
Cas des cellules
v�g�tales :
Elles sont
entour�es d�une paroi rigide qui les maintient. Lors de la turgescence, elles
n�explosent pas et ne font pas que gonfler.
� L�osmose inverse :
= Proc�d� de
filtration.
On emploie la membrane pour agir comme un filtre afin de cr�er de l�eau potable � partir de l�eau de mer.
Exemple : une membrane en
film de tri�ac�tate de cellulose ou en polyamide TFC.
Elles pr�sentent
des pores d�un diam�tre de 10�4 microns,
suffisamment petits pour filtrer les mol�cules les plus fines (bact�ries et m�me
virus)
v Diffusion facilit�e :
Les mol�cules
hydrosolubles (ions, glucide, acides amin�s) ne peuvent diffuser � travers la
membrane lipidique � des vitesses suffisantes pour satisfaire les besoins des
cellules. Le transport de ces mol�cules est assur� par un groupe de prot�ines
membranaires int�gr�es sp�cialis�es : des prot�ines transporteurs
sp�cifiques appel�es perm�ases.
La diffusion
facilit�e ne conduit qu�� un transport dans le sens �lectrochimique
d�croissant. Autrement dit : elle se fait toujours dans le sens du
gradient, cela reste un transport passif : aucune �nergie n�est n�cessaire
pour lutter contre le gradient.
Le flux net
s�arr�te lorsque son potentiel �lectrochimique est le m�me dans les 2
compartiments s�par�s par la membrane.
Les prot�ines
membranaires qui acc�l�rent le transport des solut�s et des ions sont :
-
Les transporteurs : responsables de
la diffusion facilit�e de solut�s organiques et ions inorganiques.
-
Les canaux : responsable de la diffusion
facilit�e des ions (Na+, Clˉ, K+, Ca2+)
� Caract�ristiques :
� j :
Les solut�s
diffusent beaucoup plus vite que ne le laisse pr�voir leur caract�re hydrophile
ou leur taille.
� k :
La vitesse
diffusion pr�sente une saturation

v���������������������������������������������������
����� lorsque la concentration externe du solut�
augmente.
����������������������������������������������������� dc
� l :
Le transport
peut �tre inhib� par des r�actifs analogues structuraux du solut�.
� m :
Le transport
peut aussi �tre inactiv� par des r�actifs de prot�ines.
�� Ces caract�ristiques s�expliquent par la participation au transport d�une prot�ine membranaire pr�sente dans la membrane.
� Caract�ristique
fondamentale :
La prot�ine
existe en 2 �tats conformationnels diff�rents entre lesquels elle
oscille :
-
1er �tat : le site de
fixation pour le solut� est accessible � partir d�un des 2 compartiments
aqueux.
-
2�me �tat : ce m�me site
est accessible � partir de l�autre compartiment aqueux.
Le site de
fixation pour le solut� n�est jamais accessible simultan�ment � partir des 2
compartiments aqueux. Les mol�cules de solut� sont transport�es
individuellement � la suite du changement de conformation du
transporteur ; ce qui cr�e une limitation des activit�s de transport. Un
transporteur transporte de quelques centaines � quelques milliers de mol�cules
de solut� par seconde.
� M�canisme
de diffusion facilit� par les transporteurs :
Les transports
en diffusion facilit�e par les transporteurs ne s�expriment pas en terme de
flux mais en terme de vitesse v o� l�unit� de membrane est exprim�e en poids.
Les
transporteurs sont des prot�ines membranaires dont la cha�ne traverse la
membrane plusieurs fois et qui poss�de un site de fixation sp�cifique pour un
solut�.
Exemple : La membrane des
globules rouges qui reconna�t le D_Glucose :
On observe un
�quilibre du D_Glucose de part et d�autre de la membrane. Si on ajoute du
L_Glucose (de m�me masse mol�culaire et de m�me caract�ristique hydrophile),
celui�ci traverse la membrane par diffusion passive. Ce processus va prendre
beaucoup plus de temps.
" On a ainsi
d�montr� que ce transporteur �tait sp�cifique au D_Glucose.
Le
m�canisme se fait en 3 �tapes :�����������������������
(exemple
d�un transport de l�int�rieur vers l�ext�rieur)
� j :
Il y a
reconnaissance du solut� par le transporteur sur la face int�rieure de la
membrane. Le solut� est fix� de mani�re analogue � celle d�un substrat sur une
enzyme. La fixation est caract�ris�e par une constante de dissociation : KInt
� k :
Il y a
franchissement de la membrane par le solut� � la suite d�un changement de
conformation de la prot�ine qui voit le site de fixation passer de la face
interne � la face externe.
Il s�agit de
l��tape limitante du transport : la vitesse d�pend de la constante de
vitesse qui lui est associ�e.
� l :
Le transporteur
rel�che le solut� dans le compartiment externe avec une nouvelle constante de
dissociation : KExt.
Les flux
unidirectionnels seront entrant et sortant.
Le flux net est
en fonction du gradient de concentration du solut� de part et d�autre de la
membrane.
Un m�me
transporteur peut par ailleurs fixer 1, 2 , 3 voire m�me 4 solut�s diff�rents.
Le transport peut aller soit dans le m�me sens pour tous les solut�s, soit dans
des sens diff�rents.
On parle
ainsi :����� D�uniport pour les
transporteurs fixant qu�un solut�,
De symport pour les transporteurs fixant 2 ou plusieurs solut�s dans le m�me sens,
D�antiport
pour les transporteurs fixant 2 ou plusieurs solut�s dans des sens diff�rents.
� Syst�me
de translocation :
Les transports
sont classifi�s en fonction de la stoechiom�trie du processus de transport.
� Transport uniport :
= Une seule
mol�cule est transport�e.
Les
transporteurs uniport peuvent modifier leur conformation en pr�sence (ou
absence) du ligand de sorte qu�un transport net peut s��tablir au travers de la
membrane.
Exemple : la pompe �
protons (H+)
� Transport symport :
Il est aussi
appel� Co�transport.
2 ligands
diff�rents doivent se fixer au transporteur avant d��tre tous les 2 transport�s
de fa�on simultan�e � travers de la membrane.
Exemple : le transport du
glucose au travers de la membrane en brosse des cellules intestinales.
� Transport antiport :
Le m�canisme de
transport implique un �change d�ions de part et d�autre d�une membrane. Un ion
(ou autre ligand) est �chang� au travers de la membrane pour une autre mol�cule
distincte. Ce m�canisme est appel� Ping�Pong ou Flip�Flop.
Au niveau des
symports et antiports, les transporteurs peuvent coupler les mouvements de
diff�rentes esp�ces ioniques ou des mouvements d�ions inorganiques avec des
mouvements de diff�rentes mol�cules organiques.
Exemple : � Glucose � acide amin�
������������������ Neurotransmetteurs
Le mouvement se
fait toujours dans le sens du gradient de concentration !
v Exemple
de transfert facilit� : les transporteurs du Glucose :
Le glucose est
la source principale d��nergie des Mammif�res.
Il p�n�tre
l�organisme par un transport facilit� coupl� � la diffusion facilit� du Na+. Les
transporteurs sont pr�sents dans la membrane plasmique de presque toutes les
cellules.
R�le
premier :
Il est de
permettre le transport du glucose entre le sang et le milieu intracellulaire.
L��change conduit � un transport net dans le sens entrant afin d�alimenter
les cellules en sucre.
Il existe
diff�rents transporteurs du glucose regroup�s dans une famille. Ils sont
produits par des diff�rents g�nes exprim�s de mani�re sp�cifique dans les
diff�rents tissus.
Ils sont
constitu�s par une partie hydrophobe de 12 segments transmembranaires en
h�lices a
et leurs extr�mit�s N et C terminales sont cytoplasmiques.
� La famille des transporteurs
du Glucose :
Ces
transporteurs ont des cin�tiques diff�rentes.
-
Glut 1 :���� Il
s�agit du transporteur qui permet aux cellules de r�pondre aux besoins de bases
en glucose.
-
Glut 2 :���� Il
est en tr�s grande quantit� dans les cellules b du pancr�as. Il
sont appel�s � senseur � du glucose du milieu extracellulaire (=
sensible au glucose) et sont responsables de la s�cr�tion d�insuline par ces
cellules.
Un KT �lev� pour le
transport du glucose r�pond bien � cette fonction. La vitesse du transport
varie de mani�re lin�aire avec la concentration.
-
Glut 3 :���� Il
est pr�sent dans le muscle et les tissus adipeux.
� Caract�ristiques :
Sp�cificit� : Le Pentose et
le Fructose ne franchissent pas l��pith�lium intestinal ; par contre le
Galactose et le Glucose sont transport�s.
" Une structure
pyranose comportant un carbone en position 5� est indispensable.
Inhibition : La phloridzine (par
exemple) bloque le transport du galactose et le glucose.
Influence du
m�tabolisme : L��tude de la vitesse d�absorption du glucose par
la cellule intestinale absorption d�pend de la temp�rature et da l�addition d�inhibiteurs
du m�tabolisme (exemples : 2�4 DNP ou le cyanure HCN) qui inhibe le
transport.
Effet du Na+ : Le transport du
glucose exige la pr�sence du Na+ dans le milieu
extracellulaire. Le remplacement du Na+ par le Lithium
inhibe le transport.
Exemple de l�effet
de l�insuline sur une cellule adipeuse :
 ������������������������������ ����������������������� Glucose
������������������������������ ����������������������� Glucose
Insuline
���� R�cepteur �
insuline
������������������ �� Signal activant
������ ������
les prot�ines Glut 3 :
v Autre
exemple : les porines :
Elles sont
pr�sentes sur la membrane ext�rieure des bact�ries GRAMˉ (n�gatif) et
sont tr�s perm�ables aux ions et aux petits solut�s organiques hydrophiles
(jusqu�� une masse mol�culaire d�environ 600 Da)
La diffusion facilit�e
est catalys�e par une classe de prot�ines : les Porines. Il s�agit
d�h�t�rotrim�res constitu�s de l�association de 3 cha�nes de 30 � 36 Da.
Elles sont peu
sp�cifiques.
Chez Esch�richia
coli, on trouve la OmpF et la OmpC (= Outer_membrane_protein) qui pr�sentent
une sp�cificit� pour les compos�s cationiques.
Les porines sont
les seules prot�ines organis�es en une structure enti�rement en cha�nes b dans leur
partie membranaire.
v Ph�nom�nes de transport
actifs et les diff�rents types :
Il y a une
demande d��nergie (ATP) pour effectuer un transport actif.
� Exocytose
& Endocytose :
Ils font partie
du transport v�siculaire qui permet de faire traverser la membrane aux grosses
mol�cules et les macromol�cules.
� Exocytose :
Il s�agit du
m�canisme qui permet le passage de substances de l�int�rieur de la cellule vers
l�espace extracellulaire. Il est utilis� par la cellule pour :
-
Permettre
la s�cr�tion d�hormones,
-
�mission
de neurotransmetteurs (comme l�adr�naline ou l�ac�tylcholine),
-
S�cr�tion
du mucus (pour la protection des muqueuses)
�
Processus
:
j� Avant d��tre �limin�s par la cellule, les
�l�ments destin�s � cette expulsion sont enferm�s dans un sac constitu� d�une
membrane (v�sicule)
k� Cette v�sicule migre en direction de la
membrane cytoplasmique et fusionne avec celle�ci.
l� Le contenu de la v�sicule est lib�r� dans le
liquide interstitiel. La vidange de la v�sicule ne sera obtenue que s�il y a
fusion entre la membrane de la v�sicule et celle de la cellule.
Cette fusion est
due � des prot�ines d�ancrage.
� Phagocytose :
� phagein �
= manger
Elle permet
l�internalisation de particules de grandes tailles (sup�rieure � 1 �m) par les
phagocytes : les macrophages et les neutrophiles (= forme mature de
leucocytes granuleux qui ont des propri�t�s de chimiotactisme, d�adh�rence et
de phagocytose)
Elle intervient
dans l�hom�ostasie tissulaire et assure la d�fense de l�organisme contre les
invasions en permettant l��limination des micro�organismes et la mise en place
de la r�ponse immunitaire.
La
reconnaissance de la particule d�pend de r�cepteurs sp�cifiques pr�sents � la
surface des phagocytes. Par exemple, les r�cepteurs du noyau ferreux des
immunoglobulines sont les mieux caract�ris�es.
Au cours de la
phagocytose le sac membranaire ainsi form� est appel� le phagosome. Ce dernier
fusionne avec un lysosome, une structure cellulaire renfermant des enzymes
digestives qu�elle d�verse dans le phagosome afin de d�grader le contenu.
� Pinocytose :
� pino �
= je bois
Elle consiste en
la capture et l�absorption de gouttelettes de liquides� par macrophages, par vari�t�s de globules
blancs.
Elle fait partie
d�un ph�nom�ne plus g�n�ral qui est la phagocytose : l�action de
� boire � de la cellule.
M�canisme :
-
Il
y a la mise en place, au niveau de la cellule, d�un replis de la membrane qui
vient englober une gouttelette de lipide contenue � l�ext�rieur de la cellule.
-
La
gouttelette p�n�tre la cellule � l�int�rieur d�une v�sicule pinocytaire de tr�s
petite taille.
Ce m�canisme est
employ� par les cellules intestinales pour absorber des nutriments.
� Endocytose (par r�cepteurs
interpos�s) :
Elle est tr�s
s�lective : des r�cepteurs sont employ�s par la cellule. Ce sont des
prot�ines de la membrane cytoplasmique qui vont se lier uniquement � certaines
substances (= ligands)
Les prot�ines et
les substances ainsi li�es vont p�n�trer vers l�int�rieur de la cellule en utilisant
de petites v�sicules tapiss�es de clathrine.
Les cellules
communiquent avec l�environnement par l�interm�diaire de r�cepteurs
membranaires qui reconnaissent des compos�s � l�ext�rieur de la cellule. Ces
interactions, au niveau des membranes, cr�ent un signal intracellulaire et/ou
permettent l�entr�e de macromol�cules, voire d�agents pathog�nes.
Exemple : Les r�cepteurs
de lymphocytes importants pour la r�ponse immune : les r�cepteurs �
l�antig�ne des lymphocytes T4.
Les r�cepteurs
membranaires reconnaissent leur ligand extracellulaire et cette interaction
l�endocytose de l�ensemble ligand�r�cepteur.
Les voies de
l�endocytose :
L�endocytose par
r�cepteurs permet aux cellules de communiquer avec leur environnement par
l�interm�diaire de r�cepteurs qui capturent des macromol�cules pr�sents dans le
milieu extracellulaire.
Le
m�canisme :
-
Des
r�cepteurs sont localis�s dans des r�gions particuli�res de la membrane
plasmique. Ces puits se creusent pour former des v�sicules recouvertes de
clathrine qui bourgeonnent � partir de la membrane.
-
Ensuite,
ces v�sicules perdent leur manteau de clathrine.
-
Les
v�sicules transportent les complexes r�cepteur�ligand.
-
Il
y a fusionnement avec des compartiments membranaires intracellulaires, les
endosomes.
Il existe les
outils permettant de bloquer sp�cifiquement la voie d�endocytose passant par
les puits : des mutants n�gatifs dominants de la prot�ines Eps 15.
Clathrine : une couche de
prot�ines formant la surface du cytoplasme de la v�sicule.
L�utilisation de
l�endocytose :
Elle est utilis�e par les reins pour favoriser l�absorption de diverses substances telles que le Fer et les lipoprot�ines � basse densit� comme le cholest�rol et l�insuline.
Cas
pathologique :
maladie h�r�ditaire comme l�hyper cholest�rol�mie qui est une maladie
coronarienne. Les coronaires sont les art�res irrigant le c�ur.
Le sang qui
alimente le c�ur a une concentration en cholest�rol trop importante ; ce qui
cause une baisse de la fluidit� du sang. Cela bouche les coronaires causant un
infarctus du myocarde (aussi possible avec un caillou)
Le probl�me ne
se pose pas au niveau de l�art�re principale (qui est encore grosse), mais au
niveau des art�rioles (qui sont plus fines)
Les 3 types de
l�endocytose :
Il y a une perte
d�une portion de la membrane plasmique � chaque fois qu�un sac membranaire
p�n�tre dans le cytoplasme.
L�endocytose est
si intense dans une cellule qu�une fraction de la membrane plasmique est
internalis�e chaque heure : les composants de la membrane plasmique
(prot�ines et lipides) retournent continuellement � la surface cellulaire dans
un cycle d�endocytose�exocytose (qui est assur� par les puits et les v�sicules
recouvertes de clathrine)
L�endocytose et
l�exocytose ont lieu de fa�on incessantes dans la plus part des cellules
eucaryotes. La quantit� de membrane plasmique des cellules matures varie peu �
long terme : l�ajout de membrane cons�cutif � l�exocytose compense la
perte (r�sultant de l�endocytose)
v Exemple de transport
actif : la pompe NA+�K+�ATPase :
� Mise
en �vidence :
Travaux sur
l�axone g�ant de Calmar incub� dans un liquide physiologique contenant du *Na+ (radioactif)
"� Le *Na+ p�n�tre par
diffusion passive dans l�axone en fonction du gradient �lectrochimique du *Na+.
j L�axone est
plac� dans un milieu normal : milieu s�enrichit progressivement de *Na+ qui ne peut
venir que de l�axone (sortie de Na+)
Conclusion : Le m�canisme
est un transport actif qui s�oppose au gradient �lectrochimique du Na+.
k L�axone est
empli de *Na+ et plong� dans un milieu � 0 �C
contenant du Dinitroph�nol (DNP) qui est un inhibiteur du m�tabolisme
(inhibiteur d�ATP) Le flux sortant de *Na+ est arr�t�.
L�adjonction
d�ATP au milieu entra�ne la reprise de ce flux dont l�importance est
proportionnelle � la quantit� d�ATP. L�arr�t est donc r�versible.
Conclusion : La sortie de *Na+ est donc un
ph�nom�ne actif li� � des processus m�taboliques.
l L�axone est
empli de *Na+ et plong� dans un milieu
d�pourvu de d�ions K+. le flux
sortant de *Na+ s�arr�te
�galement.
Conclusion : La sortie de *Na+ d�pend de la
pr�sence d�ions K+ dans le milieu.
m On modifie la
concentration interne en Na+.
Conclusion : La sortie de *Na+ est en fonction
de la concentration interne en Na+.
Avec ces
exp�riences, on d�crit le mod�le de la pompe Na+�K+�ATPase qui est
une prot�ine :
-
La partie ext�rieure : 3 sites de
liaisons :
-
2
sites de liaison pour les ions K+,
-
1
site de liaison pour l�ouaba�ne
(=
mol�cule digitalique bloquant la pompe)
-
La partie int�rieure (vers le cytoplasme de la
cellule) :
-
3
sites de liaison pour les ions Na+,
-
1
site de liaison de l�ATP.
" 3 ions Na+ sont expuls�s
de la cellule,
" 2 ions K+ entrent dans la
cellule.
Exemple :
Chez les
personnes �g�es, on cr�e une inhibition de la pompe Na+�K+. Cette derni�re
est remplac�e par la pompe Na+�K+. ainsi, on
obtient une entr�e de Na+ et une sortie
de Ca2+ permettant la fortification du
c�ur.
� Caract�ristiques
:
L�activit� de
cette pompe entra�ne une distribution in�gale des concentrations des ions Na+ et K+ � travers de la
membrane et donc un gradient �lectrochimique.
Chaque pompe Na+�K+�ATPase
hydrolyse 100 ATP par seconde.
1/3 de l��nergie
cellulaire est utilis�e par la pompe Na+�K+�ATPase dans les
cellules.
70 % de
l��nergie cellulaire est utilis�e par la pompe Na+�K+�ATPase dans les
cellules excitables (neurones, cellules excitables)
Il existe
d�autres pompes dans les membranes :
-
La
pompe Na+�H+,
-
La
pompe H+�K+,
-
La
pompe Na+�Ca+ et Clˉ,
-
Il
existe des inhibiteurs de la pompe Na+�H+ : les
digitaliques.
v Le co�transport :
transport par une prot�ine membranaire de 2 solut�s diff�rents :
Une pompe peut
amorcer indirectement le transport passif d�un autre solut� � travers d�une
perm�ase. C�est le m�canisme de co�transport.
Une substance
transport�e activement � travers de la membrane peut produire du travail en
diffusant en sens inverse.
� Cellules
v�g�tales :
Le gradient
�lectrochimique engendr� par la pompe � protons (par transport actif) est
utilis� pour alimenter le transport passif des acides amin�s, certains glucides
et des nutriments vers l�int�rieur de la cellule.
Ce transport
passif est assur� par une perm�ase sp�cifique poss�dant 2 sites
r�cepteurs : 1 site pour le proton et 1 site pour le saccharose. Cette
perm�ase couple donc le retour des protons au transport du saccharose.
Int�rieur������������������������������������������������������������������������������������������������� Ext�rieur
 ����������������������������������������� H+
����������������������������������������� H+
������������������������������ ������ H+
������ Saccharose���������������������������������� ���� Pompe �
protons
������������������������������ Perm�ase
����������������������������������������������������������������������������� ������ H+
( Symport )
����������������������������������������������������������������� Saccharose
Le d�placement
simultan� (co�transport) de 2 solut�s diff�rents importe le saccharose dans la
cellule � l�encontre de son gradient de concentration mais seulement s�il
� voyage � en compagnie d�un proton.
Le m�canisme
permet aux v�g�taux d�acheminer le saccharose produit par photosynth�se vers
les nervures des feuilles.
v Propri�t�s �lectriques de
la membrane neurale :
� Rappel
:
La membrane est
constitu�e d�une bicouche lipidique dans laquelle s�ins�rent des prot�ines. Les
prot�ines membranaires sont des mol�cules �lectriquement charg�es et peuvent
conduire un courant �lectrique (comme tout �l�ment porteur de charges)
Les prot�ines
sont sch�matis�es par une conductance G ou par une r�sistance R
(�gale � l�inverse de G) :������ R
=
1/G
Une conductance
varie de 10�4 � 10�8 Siemens par cm�2.
Le circuit
�lectrique �quivalent � la membrane neurale correspond donc � un g�n�rateur de
courant (une pile) de 67 mV dont le p�le + est orient� vers le
compartiment extracellulaire et le p�le � est orient� vers le
compartiment intracellulaire.
Ce g�n�rateur
alimente une r�sistance membranaire, RM, dont la valeur
est directement en fonction de la quantit� des prot�ines incluses. Dans une
portion de membrane, la quantit� de prot�ines est variable.
U = R . I
� l�inverse, la
bicouche lipidique est constitu�e de phospholipides non conducteurs, ce qui
conf�re � la membrane des propri�t�s capacitives de la membrane repr�sent�es
par un condensateur (= 2 �l�ments conducteurs s�par�s par un isolant) de
capacit� CM.
La valeur de la
capacit� membranaire CM varie peu d�un
�l�ment membranaire � l�autre.
������������������������������ En moyenne :
CM � 1 �F.cm�2
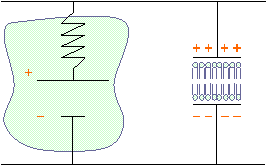
RM
����������������������������������������������������������������� CM����������������� Lipides
������ Prot�ines
On observe une
variation rapide de courant : il s�agit d�une variation de potentiel de
membrane qui suit une courbe exponentielle du fait des propri�t�s capacitives, CM, de la
membrane.
La r�sistance
transversale, RM, et la
capacit�, CM, permettent de
d�terminer la constante de temps t.����������� ����������� t�
=� RM . CM�
��� = temps
n�cessaire pour que la tension aux bornes de la membrane atteigne 63 % du
maximum.


![]()
![]() ����������������������������������������������������������������������������� ����� Charge����� ���� D�charge
����������������������������������������������������������������������������� ����� Charge����� ���� D�charge
v Propri�t�s passives de la
membrane :
On �tudie
l��volution de l�amplitude de la r�ponse �lectrotonique en injectant un courant.
Ce dernier peut �tre positif (on produit alors une d�polarisation) ou n�gatif
(on produit alors une hyperpolarisation)
En appliquant un
courant en un point A, on mesure la tension obtenue aux points B, C et D de
plus en plus �loign�s du point d�application.
" On observe que
l�amplitude de la tension va en diminuant de B � D. La tension diminue avec la
distance.
Ce courant peut
s�expliquer en raison de la r�sistance longitudinale, RL, de chaque
segment A � B, B � C et C � D� de la
fibre nerveuse. La chute de tension est �gale au produit de ce courant par la
r�sistance de ce segment :������ ����������� U = RL . I
������������������������������ I![]()
����������������������������������������������������������������� Pente� =� 1/R

![]() ����������������������������������������������������������������� ��� U
����������������������������������������������������������������� ��� U
� �volution
des tensions :
La chute de
tension suit une courbe exponentielle :������� DVX� =� DV0 . e �X/l
-
DVX : Amplitude
de la variation du potentiel enregistr�e � la distance x,
-
DV0 :
Amplitude du potentiel �lectrotonique au point d�injection du courant,
-
��x� : Distance entre
le point d�enregistrement du potentiel et le point d�injection du courant,
-
�l
:
constante d�espace. Elle est d�finie comme la distance correspondant � une
diminution de la tension de d�part de 63 %.

![]() ��� VM
��� VM
(en mV)
![]()
![]() ��
�37 %
��
�37 %
![]() ���������������������������������������������������������������������������������������� Distance
(en mm)
���������������������������������������������������������������������������������������� Distance
(en mm)
������������������ ������ l
� Signification
de la constante d�espace :
������������������������������ DVX� =� DV0 . e �X/l
Si x = 0, ���� e �X/l = 1������������������� DVX� =� DV0
Si x = l, ��� e �X/l = 0,37�������������� DVX� =� DV0 . 0,37
Les 2 milieux
intra� et extracellulaire ne sont pas des conducteurs parfaits : ils
pr�sentent une r�sistance vis���vis du passage du courant �lectrique. Elle est
sch�matis�e par une r�sistance longitudinale, RL.
� Facteurs
influen�ant la constante d�espace :
l�
=� √( RM
/
RI )
R :
r�sistance d�un segment conducteur de la membrane ayant un volume correspondant
� 1 cm de longueur et 1 cm2 de surface.
Pour une fibre
de longueur L :
������ RM� est inversement proportionnelle � la surface
de la fibre (= 2.π.R.L)
������ RI� est inversement proportionnelle au volume de
la fibre (= π.R2.L)
� R�le
de la r�sistance longitudinale au niveau de la mesure de la tension :
Si la r�sistance
est faible :
9 Faible chute de
la tension,
9 Le signal est conduit sur une
grande distance,
9 La constante
d�espace sera �lev�e.
![]()
Si la r�sistance
est forte :
9 Chute
importante de la tension,
9 Le signal est conduit sur une
petite distance,
9 La constante
d�espace sera faible.
Et
inversement
La r�sistance
longitudinale, RL, est
directement li�e au volume de l�axoplasme :
Un axone de
faible diam�tre :
9 Faible volume
de l�axoplasme,
![]() 9 Une RL importante et
une l faible.
9 Une RL importante et
une l faible.
Un axone de gros
diam�tre :
9 Volume
important de l�axoplasme,
9 Une RL faible et une
grande l.
La propagation
des ph�nom�nes �lectriques le long des fibres nerveuses sera plus ou moins
efficace selon le diam�tre de celles�ci.
La membrane des
neurones est mauvaise conductrice. Une stimulation �lectrique est d�form�e en
raison du syst�me capacitif (RM . CM) : la
stimulation ne se propage que sur de courtes distances li�es au diam�tre des
fibres nerveuses.
v Potentiel de repos :
La bicouche
lipidique �tant imperm�able aux ions, les mouvements passifs de ceux�ci ne se
font qu�au niveau des prot�ines transmembranaires sp�cialis�es, prot�ines �
canaux, dont la structure tridimensionnelle d�limite un pore aqueux � travers
duquel passent s�lectivement certains ions.
� Gen�se
des potentiels transmembranaires :
La diff�rence de
concentration d�un ion de part et d�autre de la membrane cr�e un gradient de
concentration.
Exemple : Le potassium K+ :
Chez les
Mammif�res, ����� [K+ ]INT > [K+ ]EXT
Le gradient de
concentration de part et d�autre de la membrane a tendance � faire sortir les
ions K+ vers l�ext�rieur.
Pour le gradient
�lectrique : le courant (= intensit�) va de la borne positive � la
borne n�gative. Pour le potassium K+, l�exc�s qui va
vers l�ext�rieur cr�e un gradient �lectrique allant de l�ext�rieur vers
l�int�rieur (� l�inverse du gradient de concentration)
Le passage
progressif des ions K+ (de l�int�rieur
vers l�ext�rieur) fait augmenter le gradient �lectrique jusqu�� devenir
�quivalent au gradient de concentration, mais dans le sens oppos�.
� l��quilibre,
les mouvements des ions K+ de l�int�rieur
vers l�ext�rieur sont aussi intenses que dans le sens inverse.
� Notion
de potentiel d��quilibre :
Sous l�influence
du gradient de concentration, un cation passe d�un compartiment A vers le
compartiment B, cr�ant :����������� un
exc�s de charges positives dans le compartiment B,
����������������������������������������� et un
exc�s de charges n�gatives dans le compartiment A.
Le travail W
n�cessaire pour le d�placement d�un ion d�pend :
-
De
la constante des gaz parfaits :� R
= 8,314 Joules.mol�1.K�1 ,
-
De
la temp�rature absolue :� T en
Kelvin = T en Celsius + 273 ,
-
Des
concentrations dans chaque compartiment : [cation]A ; [cation]B
W1� =� R .T . Ln([cation]A/[cation]B)
Le transfert des
cations de A vers B polarise la membrane et cr�e un gradient �lectrique de B
vers A qui tend � s�opposer au gradient de concentration.
Le travail W
s�opposant � la diffusion de l�ion d�pend :
-
De
la valence de l�ion :� z,
-
De
la quantit� d��lectricit� qui repr�sente un ion gramme :
F
= 96 500 Coulombs ,
-
De
la force �lectromotrice g�n�r�e : E
W2� =� z . F . E
Le flux ionique
n� d�un gradient de concentration est autolimit� par le gradient �lectrique
qu�il g�n�re.
B =
extracellulaire������������� A =
intracellulaire����������������� Ln = 2,3
. Log10
� l��quilibre :� ����������� z . F . E� =� R .T . Ln([cation]A/[cation]B)

���������������������������������� � �R . T����������� [cation]B
![]()
![]() EION� =� �������� �. Ln
EION� =� �������� �. Ln
�� z . F ��������� [cation]A
� 20 �C :��������������� ���������������������������������������������������������������������� 2,3
. R . T /(z . F) = 58,2
���������������������������������������������������������� [cation]B
![]() EION� =�� 58,2�
. Ln
EION� =�� 58,2�
. Ln
����������� [cation]A
En fonction des
concentrations ioniques de part et d�autre de la membrane, on peut d�terminer
le potentiel d��quilibre des ions.
-
Le
potentiel d��quilibre des ions K+ :������ EK = � 87 mV,
-
Le
potentiel d��quilibre des ions Na+ :���� ENa
= + 60 mV,
-
Le
potentiel d��quilibre des ions Ca2+ :�� ECa
= + 120 mV,
-
Le
potentiel d��quilibre des ions Clˉ :���� ECl = � 61 mV.
(valeurs
moyennes pour les cellules de Mammif�res)
Pour effectuer
ces calculs, il faut partir du principe que la membrane est perm�able � un seul
ion et imperm�able aux autres. Ces valeurs peuvent varier, selon les cellules,
en fonction des concentrations ioniques des milieux intra� et extracellulaire
et fonction de la temp�rature de l�esp�ce � laquelle appartiennent les cellules
�tudi�es.
� Du
potentiel d��quilibre au potentiel de membrane :
Le potentiel de
membrane n�est pas seulement un compromis entre les potentiels d��quilibre. Il d�pend
�galement de la pompe �lectrog�nique Na � K et du gradient �lectrochimique qui
conditionne la force de diffusion (ou � driving force �)
� Potentiel de membrane :
�quation de Goldman�Hodgkin�Katz :
= �quation du
champ constant.
-
Le
champ �lectrique de la membrane est constant.
-
Les
ions se d�place ind�pendamment les uns aux autres.
-
Chaque
ion poss�de un coefficient de perm�abilit�.
Coefficient
de diffusion
PIon�� =�� bIon� .� DIon� / DX
Perm�abilit�
� Coefficient����������������� �paisseur
����������������������������������������� de
partage������������������������������ de
la membrane

����������������������������������������������������� R
. T�������������� PK.[K+]Ext + PNa.[Na+]Ext + PCl.[Clˉ]Int
![]()
![]() Potentiel de
membrane :� VM� =� ����������� .
�Ln
Potentiel de
membrane :� VM� =� ����������� .
�Ln
����������������������������������������������������� z . F�������������� PK.[K+]Int + PNa.[Na+]Int + PCl.[Clˉ]Ext
� Il faut faire attention � la valence de
l�ion Clˉ qui est n�gative.
Les ions qui
sont distribu�s passivement (pas par transport actif) ne peuvent d�terminer le
potentiel de repos. Le Clore, Clˉ, est distribu� passivement : on ne
peut pas en tenir compte dans l��quation.

������������������������������ ����������� ��
PK.[K+]Ext + PNa.[Na+]Ext
![]() EM� =� � 61 mV
. Log
EM� =� � 61 mV
. Log
������������������������������ ����������� ��
PK.[K+]Int + PNa.[Na+]Int
En connaissant
les concentrations ioniques, on peut calculer PNa /PK. Le potentiel
de repos sera d�terminer par ce rapport.
Le Calcium Ca2+ n�a pas �t�
pris en compte car il a un effet n�gligeable sur le potentiel de repos d�un
neurone. Les concentrations [Ca2+] aux niveaux
intra� et extracellulaire sont faibles. Ce ne serait pas le m�me cas s�il on
�tudiait des cellules cardiaques.
Pour le neurone,
la formule est simplifi�e du fait que le Sodium et Potassium aient la m�me
valence +1.
� Conduction ionique
membranaire :
Pour un canal
ionique, la conductance caract�rise la facilit�e avec laquelle les ions
traversent le pore aqueux de la prot�ine � canal.
" La conductance
membranaire d�une cellule pour un ion (= conductance ionique membranaire) gIon (g)Ion est proportionnelle :
-
�
la conductance �l�mentaire d�un canal ionique gIon,
-
Au
nombre total de canaux de l�esp�ce ionique dans la membrane NIon,
-
�
la probabilit� P0 que ces canaux
soient � l��tat ouvert.
gIon (g)Ion� = �gIon . NIon . PIon
gIon varie entre 10 et 200 pS (= 10�12 S) selon du
type de canal.
" Mesure des
courants ioniques.
L�intensit� des
courants ioniques traversant un canal (appel� courant ionique �l�mentaire) est
�gal � :�����������
������������������������������ IIon� =� gIon (VM � EIon)
� Potentiel
de repos de la membrane neuronale :
Il s�agit d�une
cellule dont la membrane comporte des canaux K+ et Na+ ouverts. Elle
dispose de transports actifs qui permettent de maintenir les concentrations des
ions K+ et Na+ de part et
d�autre de la membrane constantes.
" Les potentiels
d��quilibre des ions K+ (EK) et Na+ (ENa) restent constants.
Pour les
neurones, on n�utilise que le K+ et Na+.
Au potentiel de
repos, le potentiel de membrane, VM, atteint un
�tat stable quand le flux net de charges (K+ et Na+) sera nul en
terme de courant ionique :
INa�
+� IK� =� 0
Avec les
�quations pr�c�dentes :�������������� IK� =� gK (VM � EK)
����������������������������������������� ����������������������� INa� =� gNa (VM � ENa)
"������� gK (VM � EK)�
+� gNa (VM � ENa)� =� 0
������������������ VM� =� (gK . EK� +� gNa . ENa) / (gK + gNa)
La valeur du potentiel de membrane d�pend de la conductance relative des 2 ions (leurs potentiels restent constants) et du rapport gK /gNa.
Si� a = gK /gNa����������������������� "������� VM� = �( EK�
+� a . ENa) / a
Exemple de la
membrane neuronale :
VM� =� � 60 mV��������������������������������� EK� =� � 87
mV���������� ENa� =� + 60 mV
"� a = 0,2������������� La conductance des ions K+ est 5 fois plus
grande que celle des ions Na+.
������������������ a �= �gK �=
�5 gNa
La cellule au
repos : il y a beaucoup plus de canaux K+ ouverts que de
canaux Na+.
En r�alit�, il
n�y a pas une diff�rence de 5 entre les 2 ions car le gradient �lectrochimique
de K+, (VM � EK) = +
27 mV,
est moins important que le gradient �lectrochimique de Na+, (VM � ENa) = � 120 mV.
En r�alit�, 2
ions K+ sortent de la cellule quand 3
ions Na+ entrent.
v Gradient
�lectrochimique :
Pour
traverser la membrane, un ion est soumis � un gradient �lectrochimique, appel�
aussi � driving force �, qui s�exprime par la diff�rence entre le
potentiel de membrane et le potentiel d��quilibre de l�ion. Cela cr�e un flux
net proportionnel � ce gradient.������������� VM� = � 60 mV
|
Potentiel
d��quilibre de l�ion : |
Gradient
�lectrochimique : |
Flux
net : |
|
EK� =� �
87 mV���������� |
(VM � EK) = + 27 mV |
Sortant |
|
ENa�
=� + 60 mV |
(VM � ENa) = � 120 mV |
Entrant |
|
ECa�
=� + 120 mV |
(VM � ECa) = � 180 mV |
Entrant |
|
ECl�
=� � 61 mV |
(VM � ENa) = + 1 mV |
�quilibre |
Par
convention :
������ Le flux net est positif quand un cation a
tendance � sortir de la cellule.
������ Le flux net est n�gatif quand un cation a
tendance � entrer de la cellule.
� �volution
du potentiel de membrane :
Au cours d�une
d�polarisation cellulaire, il y a une entr� de Na+ et une sortie
de K+.
Derni�re page du
3�me poly (blabla de r�sum�)
Il s�agit d�un
ph�nom�ne �lectrique qui pr�sente 2 propri�t�s remarquables :
-
Il
se d�veloppe d�une mani�re � tout ou rien �.
-
Lorsqu�il
est �mis en un point, il se propage sans att�nuation.
v Sch�ma d�un potentiel
d�action :
Il se d�roule en
3 �tapes :
� 1�re
�tape : la phase de mont�e :
Ou phase
ascendante. Elle consiste en une d�polarisation tr�s rapide qui atteint sa
valeur maximale en moins d�une milliseconde. L�amplitude maximale est voisine
de 100 mV.
M�canisme :
Il y a une
ouverture des canaux Na+ qui sont
sensibles � la d�polarisation membranaire.
Au repos, � �50
mV, la probabilit� pour que les canaux Na+ soient ouverts
est tr�s faible. Les canaux restent ferm�s jusqu�� une certaine valeur de
potentiel (�
�40 mV)
" On parle de
canaux voltage d�pendants.
� partir de
cette valeur, il y a une ouverture rapide des canaux Na+ permettant
l�entr�e d�ions dans la cellule et causant la d�polarisation.
� 2�me
�tape : la phase de descente :
Ou phase descendante.
Il s�agit du processus de repolarisation, �galement tr�s rapide (1 � 2 ms),
lors duquel le potentiel de membrane revient � sa valeur initiale (potentiel de
repos)
2 facteurs
limitent la dur�e du potentiel d�action :
-
La
d�polarisation qui inactive graduellement les canaux Na+,
-
L�existence
d�un d�lai d�ouverture des canaux K+.
M�canisme :
Une fois
ouverts, les canaux Na+ ne restent pas
dans cet �tat mais s�inactivent.
Ce processus
suffirait � induire le processus de repolarisation membranaire, c�est le cas
des potentiels d�action des fibres my�linis�es.
On enregistre un
potentiel d�action dans un milieu contenant
-
Soit
du TEA (T�tra_ �thyl_Ammonium) bloquant les canaux de K+ dans leur �tat
ferm�,
-
Soit
de la pronase bloquant l�inactivit�.
On observe une
dur�e plus longue du potentiel d�action (la repolarisation est plus lente)
La
d�polarisation membranaire entra�ne l�augmentation la probabilit� d�ouverture
des canaux K+.
Ces derniers
s�ouvrent avec un d�lai et s�inactivent tr�s lentement, contrairement aux
canaux de Na+.
� 3�me
�tape : la phase de d�hyperpolarisation :
Ou
post�d�polarisation (ou after�depolarization) � la fin de la repolarisation, le
potentiel de membrane atteint bri�vement une valeur plus n�gative que le
potentiel de repos.
D :
Elle n�est pas pr�sente dans toutes les cellules excitatrices, comme chez les
cellules cardiaques.
M�canisme :
Elle est due au fait
que les canaux K+ s�ouvrent
lentement. La probabilit� que ceux�ci soient ouverts est �lev�e.
" La sortie
d�ions K+ hors de la cellule pousse � la
repolarisation suivie d�une hyperpolarisation.
= Efflux d�ions K+
hyperpolarisant.
� Circuit
�lectrique �quivalent :
En tenant compte
de la capacit� de membrane, CM, et de la
conductance membranaire, gM, ainsi que des conductances mises en
jeu lors du potentiel d�action gK et gNa :
" Le courant
traversant chaque type de canal sensible au potentiel d�action peut �tre
calcul� :���������������������� IK� =� gK (VM � EK)
������ ����������������������� INa� =� gNa (VM � ENa)
v Propri�t�s du potentiel
d�action :
Il faut que la
d�polarisation initiale atteigne une valeur minimale pour qu�il y ait �mission
d�un potentiel d�action. Cette valeur, appel�e valeur seuil, peut �tre atteinte
� 2 niveaux diff�rents.
-
Au niveau des synapses excitatrices :
La valeur seuil
est atteinte au niveau des neurom�diateurs pr�synaptiques causant la
d�polarisation dans les �l�ments post�synaptiques.
Exemple :
Aux jonctions
neuromusculaires, il y a une lib�ration d�ACh (Ac�tylcholine) par les
terminaisons pr�synaptiques.
L�ac�tylcholine
se lie � des r�cepteurs post�synaptiques et entra�ne l�ouverture de canaux
cationiques et donc une d�polarisation locale, appel�e potentiel de post�synapse
excitatrice (PPSE)
On le trouve au
niveau des synapses excitatrices.
-
Au niveau des r�cepteurs sensoriels :
Un stimulus
ext�rieur va entra�ner une variation de potentiel local, appel� potentiel de
r�ception.
� Mise
en �vidence du potentiel seuil :
On applique des
sauts de courant d�polarisants d�intensit� de plus en plus grande et on
enregistre les potentiels �lectrochimiques correspondants.
" L�amplitude
augmente avec l�augmentation du courant. � une certaine valeur de courant, on
observe le d�clenchement d�un potentiel d�action.
� P�riodes
r�fractaires absolue et relative :
Un second
stimulus, appliqu� artificiellement dans le but de d�clencher un nouveau
potentiel d�action (alors qu�il y en a d�j� un en cours), est appel�
extra�stimulus
� extra �
= en plus
Pendant environ
1 milliseconde apr�s le sommet du potentiel d�action, il est impossible de
d�clencher un nouveau potentiel d�action � chevauchant de pr�s � le
premier.
" Il s�agit de la
p�riode r�fractaire absolue : aucune d�polarisation (= stimulus) ne
peut provoquer une seconde r�ponse r�g�n�ratrice. Cette p�riode dure jusqu�� ce
que la phase descendante du potentiel d�action atteigne la valeur seuil.
Apr�s cette p�riode, il existe une p�riode r�fractaire dite relative pendant laquelle une seconde polarisation peut engendrer un nouveau potentiel d�action m�me si le premier n�est pas encore fini.
Pendant cette p�riode, la probabilit� est grande pour que les canaux K+ soient ouverts. Cette probabilit� atteint son maximum � peu pr�s lorsque le potentiel de membrane atteint la valeur seuil.
La p�riode r�fractaire relative commence � partir de ce moment.
" Lors de cette p�riode, la probabilit� que les canaux K+ soient ouverts est alors inf�rieure � celle au moment o� le potentiel avait atteint� la valeur seuil.
� Influx
nerveux :
Il s�agit du
message qui circule le long de la fibre nerveuse de l�axone et qui se
caract�rise par une succession de potentiels d�action.